

A gatekeeper chaperone complex directs translocator secretion during type three secretion
- PMID: 25375170
- PMCID: PMC4222845
- DOI: 10.1371/journal.ppat.1004498
A gatekeeper chaperone complex directs translocator secretion during type three secretion
Abstract
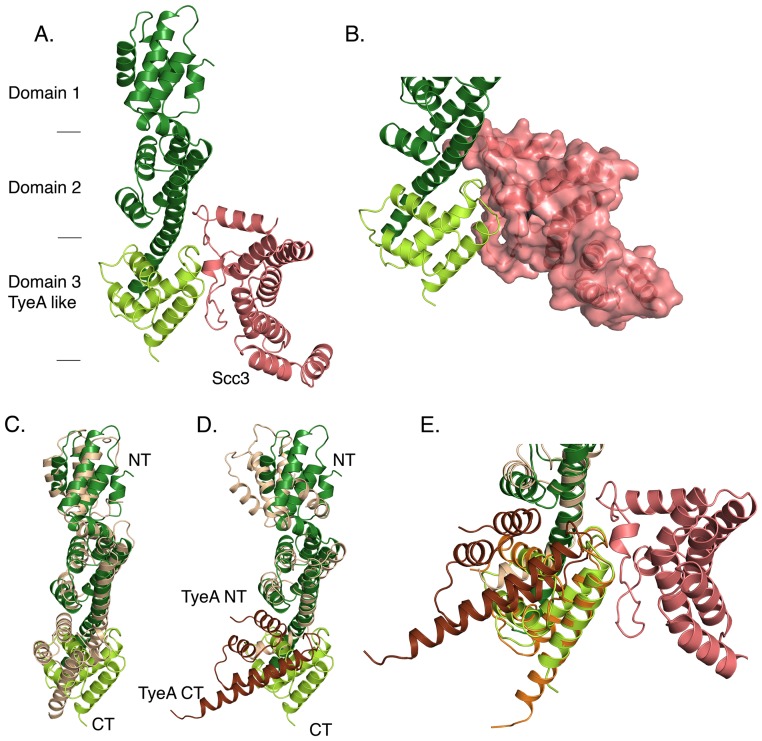
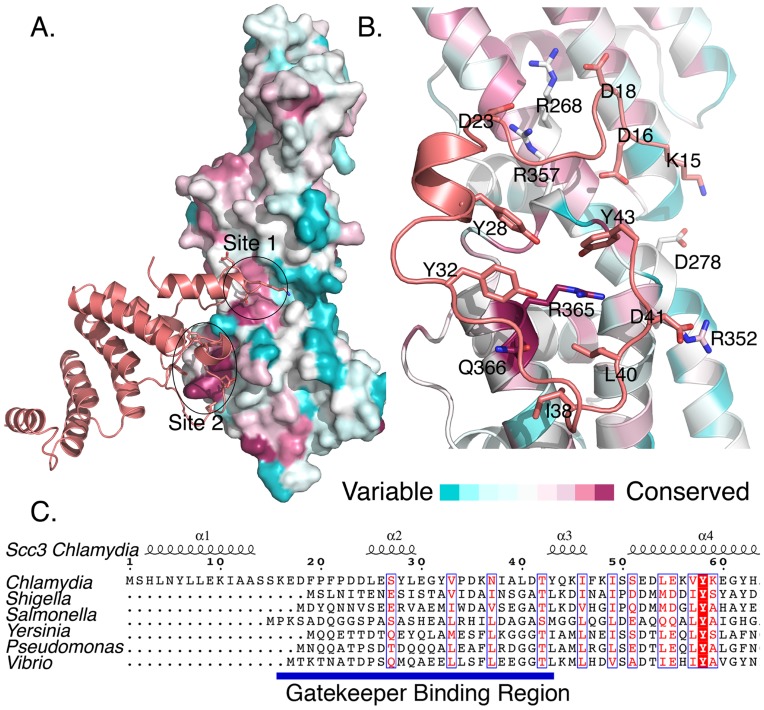
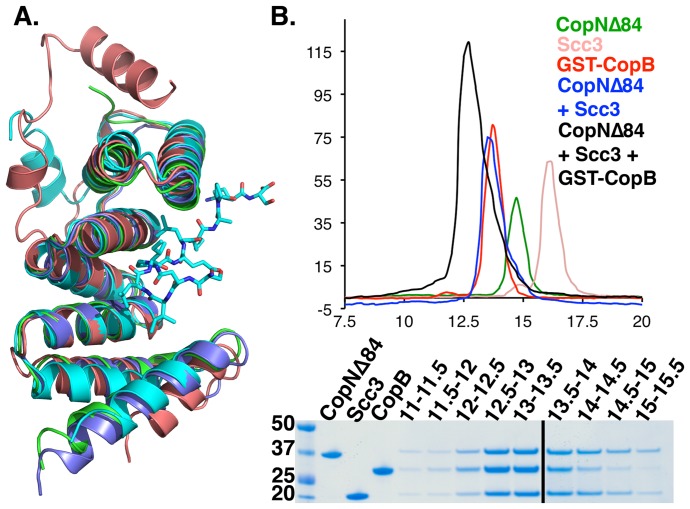
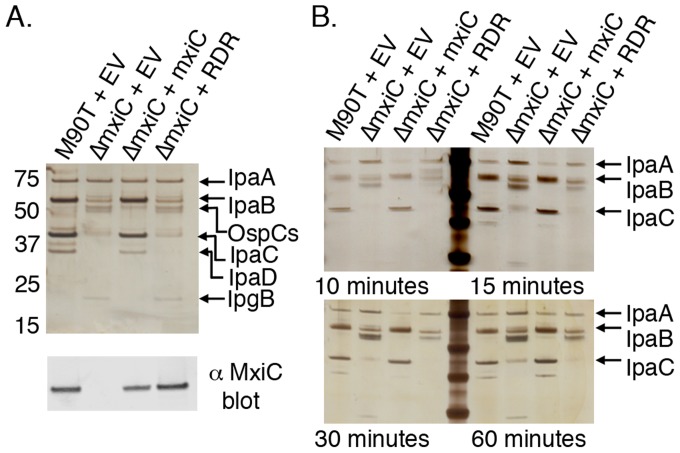
Many Gram-negative bacteria use Type Three Secretion Systems (T3SS) to deliver effector proteins into host cells. These protein delivery machines are composed of cytosolic components that recognize substrates and generate the force needed for translocation, the secretion conduit, formed by a needle complex and associated membrane spanning basal body, and translocators that form the pore in the target cell. A defined order of secretion in which needle component proteins are secreted first, followed by translocators, and finally effectors, is necessary for this system to be effective. While the secreted effectors vary significantly between organisms, the ∼20 individual protein components that form the T3SS are conserved in many pathogenic bacteria. One such conserved protein, referred to as either a plug or gatekeeper, is necessary to prevent unregulated effector release and to allow efficient translocator secretion. The mechanism by which translocator secretion is promoted while effector release is inhibited by gatekeepers is unknown. We present the structure of the Chlamydial gatekeeper, CopN, bound to a translocator-specific chaperone. The structure identifies a previously unknown interface between gatekeepers and translocator chaperones and reveals that in the gatekeeper-chaperone complex the canonical translocator-binding groove is free to bind translocators. Structure-based mutagenesis of the homologous complex in Shigella reveals that the gatekeeper-chaperone-translocator complex is essential for translocator secretion and for the ordered secretion of translocators prior to effectors.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Crystal structure of the Yersinia enterocolitica type III secretion chaperone SycD in complex with a peptide of the minor translocator YopD.BMC Struct Biol. 2012 Jun 18;12:13. doi: 10.1186/1472-6807-12-13. BMC Struct Biol. 2012. PMID: 22708907 Free PMC article.
-
A new means to identify type 3 secreted effectors: functionally interchangeable class IB chaperones recognize a conserved sequence.mBio. 2012 Feb 14;3(1):e00243-11. doi: 10.1128/mBio.00243-11. Print 2012. mBio. 2012. PMID: 22334517 Free PMC article.
-
Binding mode analysis of a major T3SS translocator protein PopB with its chaperone PcrH from Pseudomonas aeruginosa.Proteins. 2014 Dec;82(12):3273-85. doi: 10.1002/prot.24666. Epub 2014 Oct 21. Proteins. 2014. PMID: 25116453
-
Essential functions of chaperones and adaptors of protein secretion systems in Gram-negative bacteria.FEBS J. 2022 Aug;289(16):4704-4717. doi: 10.1111/febs.16056. Epub 2021 Jun 23. FEBS J. 2022. PMID: 34092034 Review.
-
Structure and biophysics of type III secretion in bacteria.Biochemistry. 2013 Apr 16;52(15):2508-17. doi: 10.1021/bi400160a. Epub 2013 Apr 5. Biochemistry. 2013. PMID: 23521714 Free PMC article. Review.
Cited by
-
Type Three Secretion System in Attaching and Effacing Pathogens.Front Cell Infect Microbiol. 2016 Oct 21;6:129. doi: 10.3389/fcimb.2016.00129. eCollection 2016. Front Cell Infect Microbiol. 2016. PMID: 27818950 Free PMC article. Review.
-
Insight into microtubule nucleation from tubulin-capping proteins.Proc Natl Acad Sci U S A. 2019 May 14;116(20):9859-9864. doi: 10.1073/pnas.1813559116. Epub 2019 Apr 29. Proc Natl Acad Sci U S A. 2019. PMID: 31036638 Free PMC article.
-
Structural and Functional Analysis of SsaV Cytoplasmic Domain and Variable Linker States in the Context of the InvA-SsaV Chimeric Protein.Microbiol Spectr. 2021 Dec 22;9(3):e0125121. doi: 10.1128/Spectrum.01251-21. Epub 2021 Dec 1. Microbiol Spectr. 2021. PMID: 34851139 Free PMC article.
-
Structural analysis of SepL, an enteropathogenic Escherichia coli type III secretion-system gatekeeper protein.Acta Crystallogr F Struct Biol Commun. 2015 Oct;71(Pt 10):1300-8. doi: 10.1107/S2053230X15016064. Epub 2015 Sep 23. Acta Crystallogr F Struct Biol Commun. 2015. PMID: 26457522 Free PMC article.
-
α-Helices in the Type III Secretion Effectors: A Prevalent Feature with Versatile Roles.Int J Mol Sci. 2021 May 21;22(11):5412. doi: 10.3390/ijms22115412. Int J Mol Sci. 2021. PMID: 34063760 Free PMC article. Review.
References
-
- Cornelis GR (2006) The type III secretion injectisome. Nat Rev Microbiol 4: 811–825. - PubMed
-
- Galan JE, Wolf-Watz H (2006) Protein delivery into eukaryotic cells by type III secretion machines. Nature 444: 567–573. - PubMed
-
- Galan JE, Cossart P (2005) Host-pathogen interactions: a diversity of themes, a variety of molecular machines. Curr Opin Microbiol 8: 1–3. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
