

Host-specific parvovirus evolution in nature is recapitulated by in vitro adaptation to different carnivore species
- PMID: 25375184
- PMCID: PMC4223063
- DOI: 10.1371/journal.ppat.1004475
Host-specific parvovirus evolution in nature is recapitulated by in vitro adaptation to different carnivore species
Erratum in
- PLoS Pathog. 2014 Dec;10(12):e1004586
Abstract
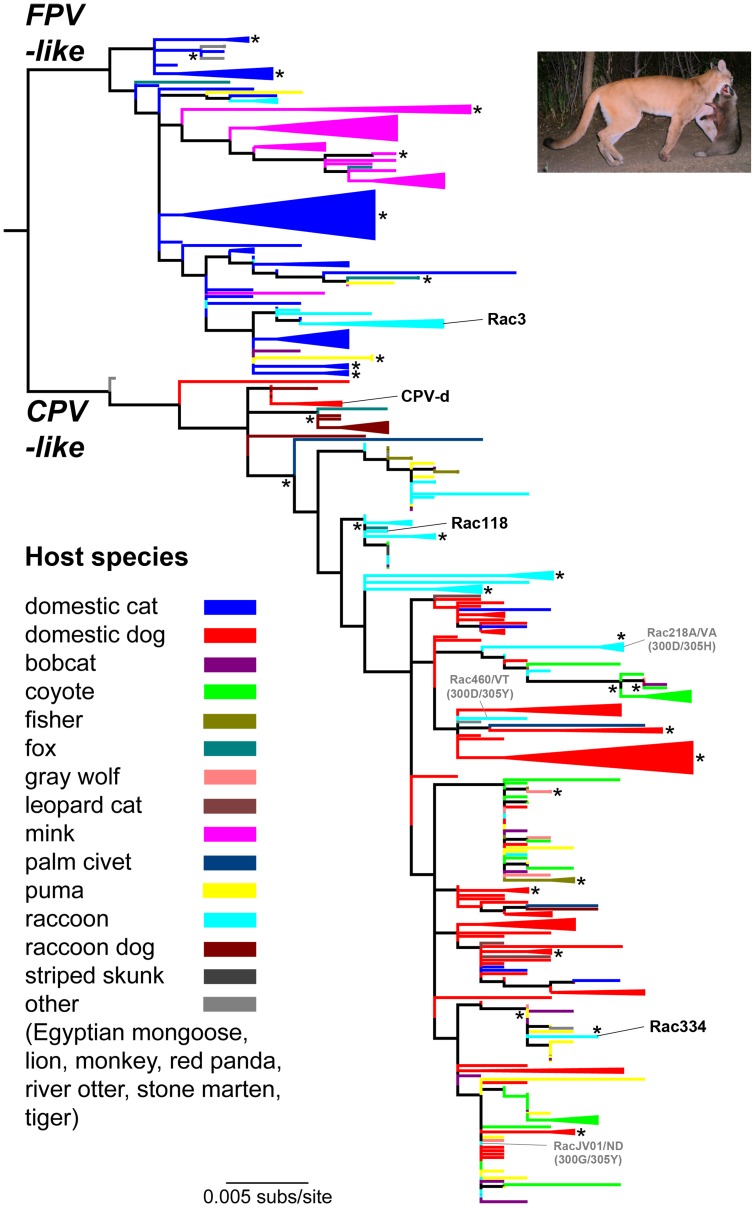
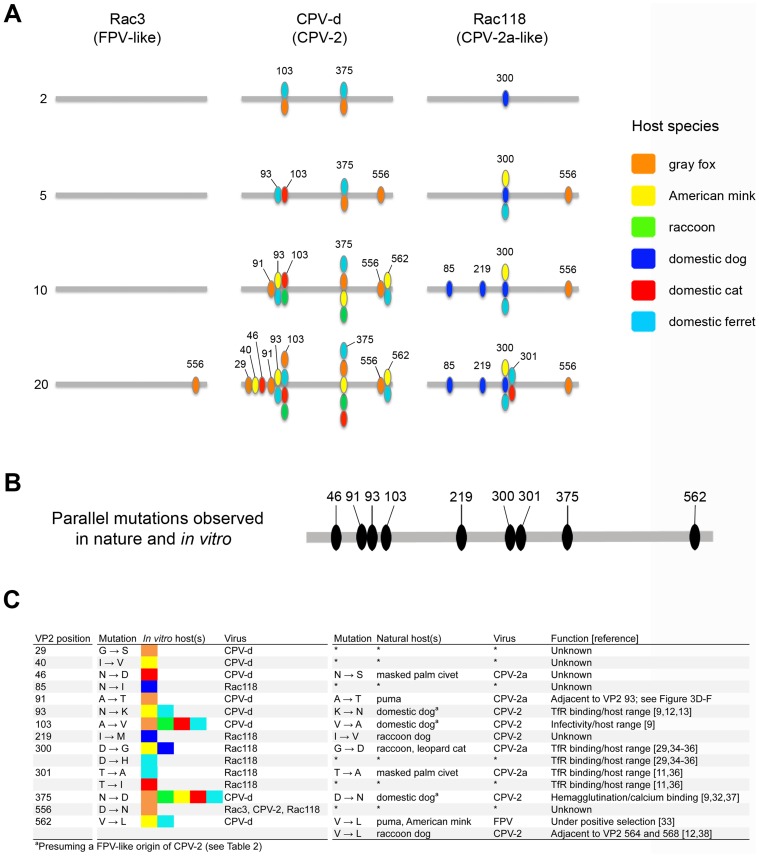
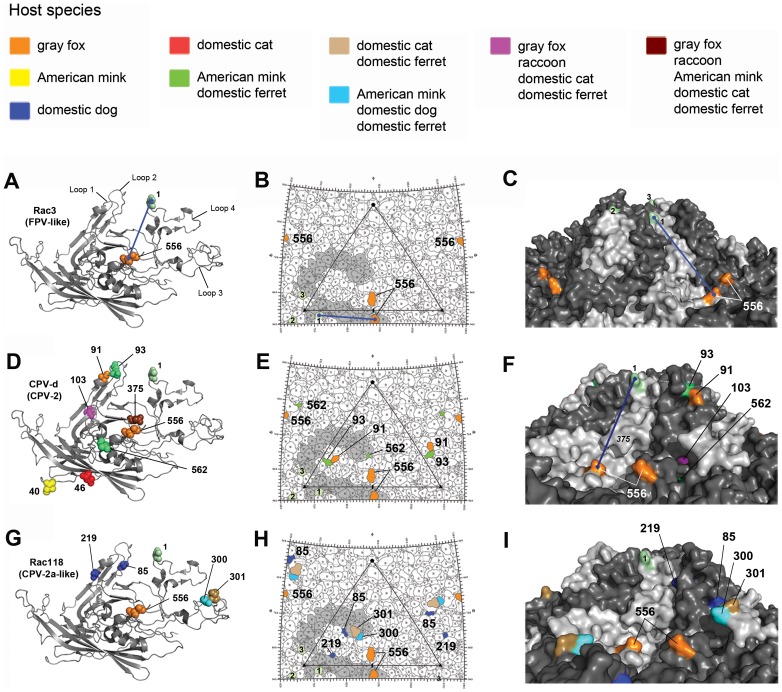
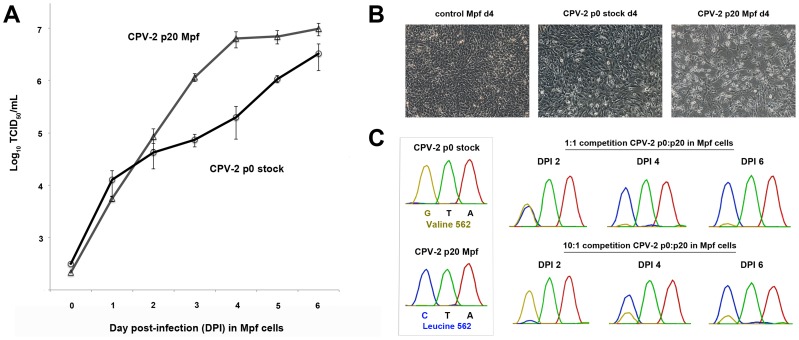
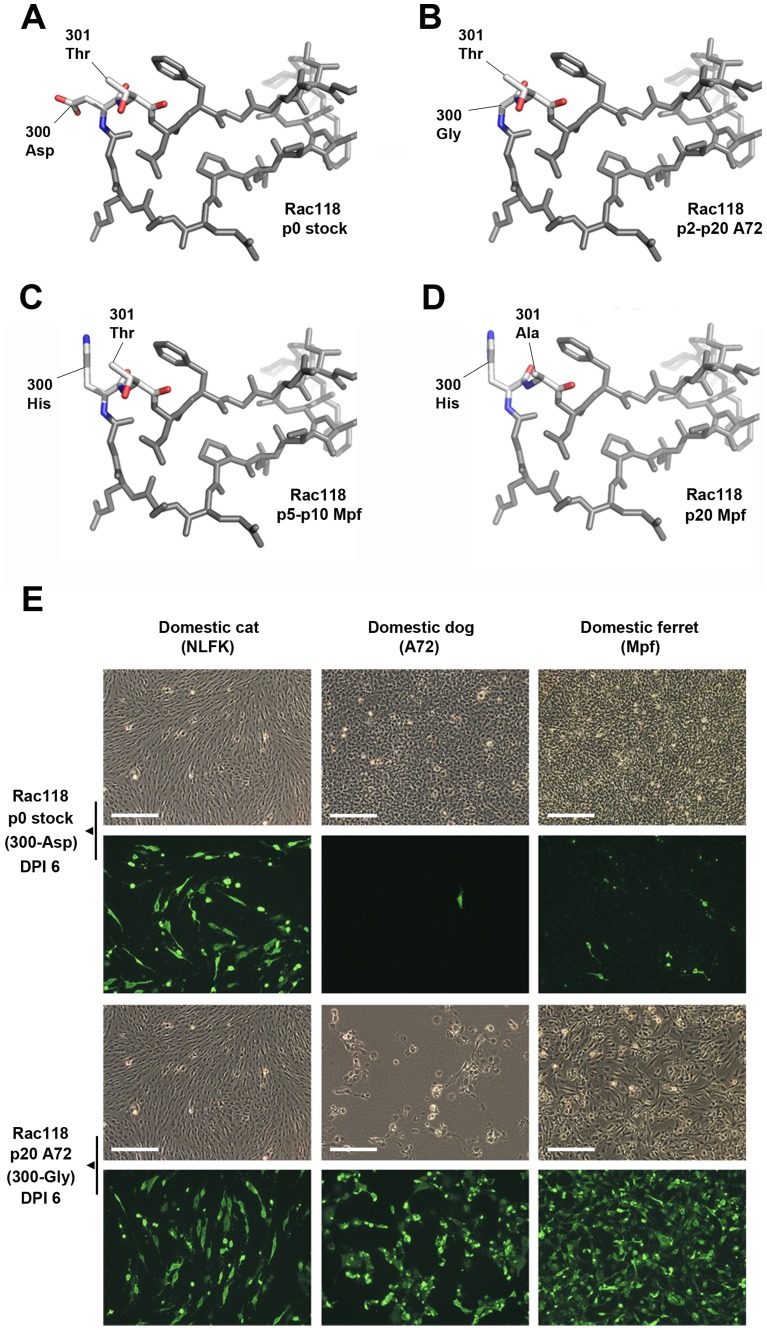
Canine parvovirus (CPV) emerged as a new pandemic pathogen of dogs in the 1970s and is closely related to feline panleukopenia virus (FPV), a parvovirus of cats and related carnivores. Although both viruses have wide host ranges, analysis of viral sequences recovered from different wild carnivore species, as shown here, demonstrated that>95% were derived from CPV-like viruses, suggesting that CPV is dominant in sylvatic cycles. Many viral sequences showed host-specific mutations in their capsid proteins, which were often close to sites known to control binding to the transferrin receptor (TfR), the host receptor for these carnivore parvoviruses, and which exhibited frequent parallel evolution. To further examine the process of host adaptation, we passaged parvoviruses with alternative backgrounds in cells from different carnivore hosts. Specific mutations were selected in several viruses and these differed depending on both the background of the virus and the host cells in which they were passaged. Strikingly, these in vitro mutations recapitulated many specific changes seen in viruses from natural populations, strongly suggesting they are host adaptive, and which were shown to result in fitness advantages over their parental virus. Comparison of the sequences of the transferrin receptors of the different carnivore species demonstrated that many mutations occurred in and around the apical domain where the virus binds, indicating that viral variants were likely selected through their fit to receptor structures. Some of the viruses accumulated high levels of variation upon passage in alternative hosts, while others could infect multiple different hosts with no or only a few additional mutations. Overall, these studies demonstrate that the evolutionary history of a virus, including how long it has been circulating and in which hosts, as well as its phylogenetic background, has a profound effect on determining viral host range.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Parrish CR, Kawaoka Y (2005) The origins of new pandemic viruses: the acquisition of new host ranges by canine parvovirus and influenza A viruses. Annu Rev Microbiol 59: 553–586. - PubMed
-
- Kuiken T, Holmes EC, McCauley J, Rimmelzwaan GF, Williams CS, et al. (2006) Host species barriers to influenza virus infections. Science 312: 394–397. - PubMed
-
- Parrish CR (1999) Host range relationships and the evolution of canine parvovirus. Vet Microbiol 69: 29–40. - PubMed
-
- Parrish CR, O'Connell PH, Evermann JF, Carmichael LE (1985) Natural variation of canine parvovirus. Science 230: 1046–1048. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
