

Regulation of mRNA abundance by polypyrimidine tract-binding protein-controlled alternate 5' splice site choice
- PMID: 25375251
- PMCID: PMC4222953
- DOI: 10.1371/journal.pgen.1004771
Regulation of mRNA abundance by polypyrimidine tract-binding protein-controlled alternate 5' splice site choice
Abstract
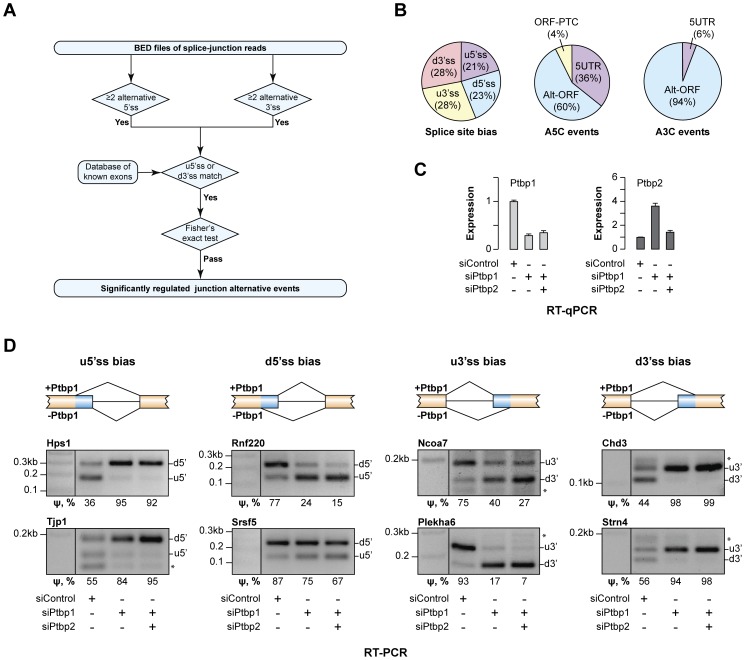
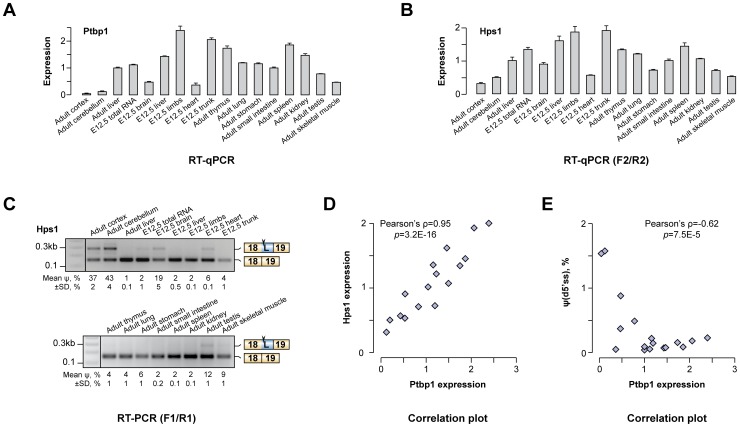
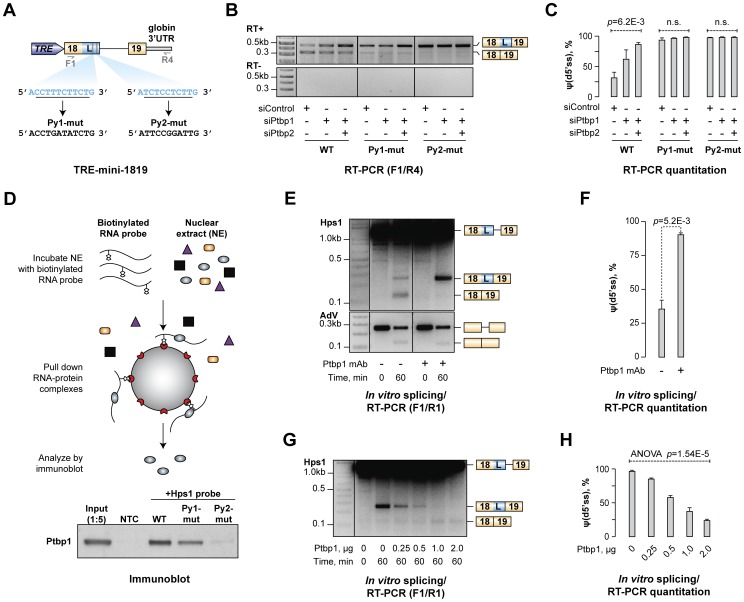
Alternative splicing (AS) provides a potent mechanism for increasing protein diversity and modulating gene expression levels. How alternate splice sites are selected by the splicing machinery and how AS is integrated into gene regulation networks remain important questions of eukaryotic biology. Here we report that polypyrimidine tract-binding protein 1 (Ptbp1/PTB/hnRNP-I) controls alternate 5' and 3' splice site (5'ss and 3'ss) usage in a large set of mammalian transcripts. A top scoring event identified by our analysis was the choice between competing upstream and downstream 5'ss (u5'ss and d5'ss) in the exon 18 of the Hps1 gene. Hps1 is essential for proper biogenesis of lysosome-related organelles and loss of its function leads to a disease called type 1 Hermansky-Pudlak Syndrome (HPS). We show that Ptbp1 promotes preferential utilization of the u5'ss giving rise to stable mRNAs encoding a full-length Hps1 protein, whereas bias towards d5'ss triggered by Ptbp1 down-regulation generates transcripts susceptible to nonsense-mediated decay (NMD). We further demonstrate that Ptbp1 binds to pyrimidine-rich sequences between the u5'ss and d5'ss and activates the former site rather than repressing the latter. Consistent with this mechanism, u5'ss is intrinsically weaker than d5'ss, with a similar tendency observed for other genes with Ptbp1-induced u5'ss bias. Interestingly, the brain-enriched Ptbp1 paralog Ptbp2/nPTB/brPTB stimulated the u5'ss utilization but with a considerably lower efficiency than Ptbp1. This may account for the tight correlation between Hps1 with Ptbp1 expression levels observed across mammalian tissues. More generally, these data expand our understanding of AS regulation and uncover a post-transcriptional strategy ensuring co-expression of a subordinate gene with its master regulator through an AS-NMD tracking mechanism.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Regulation of BCL-X splicing reveals a role for the polypyrimidine tract binding protein (PTBP1/hnRNP I) in alternative 5' splice site selection.Nucleic Acids Res. 2014 Oct 29;42(19):12070-81. doi: 10.1093/nar/gku922. Epub 2014 Oct 7. Nucleic Acids Res. 2014. PMID: 25294838 Free PMC article.
-
Crossregulation and functional redundancy between the splicing regulator PTB and its paralogs nPTB and ROD1.Mol Cell. 2007 Aug 3;27(3):420-34. doi: 10.1016/j.molcel.2007.06.016. Mol Cell. 2007. PMID: 17679092 Free PMC article.
-
A post-transcriptional regulatory switch in polypyrimidine tract-binding proteins reprograms alternative splicing in developing neurons.Genes Dev. 2007 Jul 1;21(13):1636-52. doi: 10.1101/gad.1558107. Genes Dev. 2007. PMID: 17606642 Free PMC article.
-
Regulation of alternative splicing by PTB and associated factors.Biochem Soc Trans. 2005 Jun;33(Pt 3):457-60. doi: 10.1042/BST0330457. Biochem Soc Trans. 2005. PMID: 15916540 Review.
-
PTB: Not just a polypyrimidine tract-binding protein.J Cell Physiol. 2022 May;237(5):2357-2373. doi: 10.1002/jcp.30716. Epub 2022 Mar 14. J Cell Physiol. 2022. PMID: 35288937 Review.
Cited by
-
Tissue-specific regulation of gene expression via unproductive splicing.Nucleic Acids Res. 2023 Apr 24;51(7):3055-3066. doi: 10.1093/nar/gkad161. Nucleic Acids Res. 2023. PMID: 36912101 Free PMC article.
-
Alternative splicing in EMT and TGF-β signaling during cancer progression.Semin Cancer Biol. 2024 Jun;101:1-11. doi: 10.1016/j.semcancer.2024.04.001. Epub 2024 Apr 15. Semin Cancer Biol. 2024. PMID: 38614376 Free PMC article. Review.
-
RNA Binding Protein Ptbp2 Is Essential for Male Germ Cell Development.Mol Cell Biol. 2015 Dec;35(23):4030-42. doi: 10.1128/MCB.00676-15. Epub 2015 Sep 21. Mol Cell Biol. 2015. PMID: 26391954 Free PMC article.
-
The leader RNA of SARS-CoV-2 sequesters polypyrimidine tract binding protein (PTBP1) and influences pre-mRNA splicing in infected cells.Virology. 2024 Apr;592:109986. doi: 10.1016/j.virol.2024.109986. Epub 2024 Jan 6. Virology. 2024. PMID: 38290414 Free PMC article.
-
Deep sequencing of pre-translational mRNPs reveals hidden flux through evolutionarily conserved alternative splicing nonsense-mediated decay pathways.Genome Biol. 2021 May 3;22(1):132. doi: 10.1186/s13059-021-02309-y. Genome Biol. 2021. PMID: 33941243 Free PMC article.
References
-
- Black DL (2003) Mechanisms of alternative pre-messenger RNA splicing. Annu Rev Biochem 72: 291–336. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
