

Description of Caenorhabditis sinica sp. n. (Nematoda: Rhabditidae), a nematode species used in comparative biology for C. elegans
- PMID: 25375770
- PMCID: PMC4222906
- DOI: 10.1371/journal.pone.0110957
Description of Caenorhabditis sinica sp. n. (Nematoda: Rhabditidae), a nematode species used in comparative biology for C. elegans
Abstract
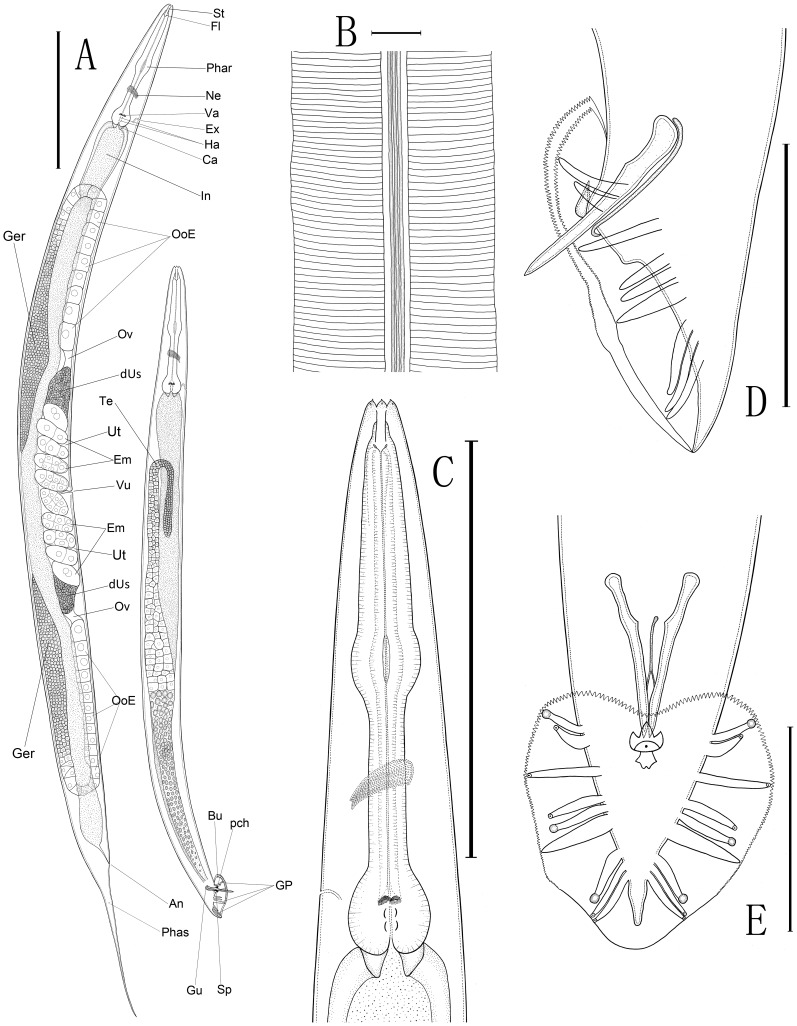
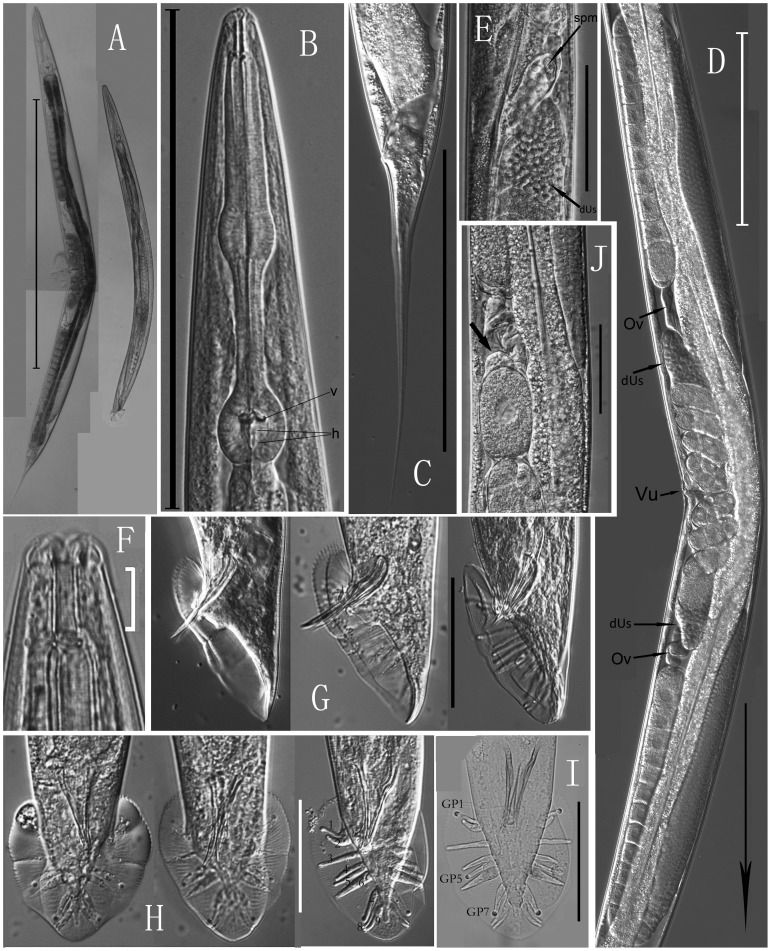
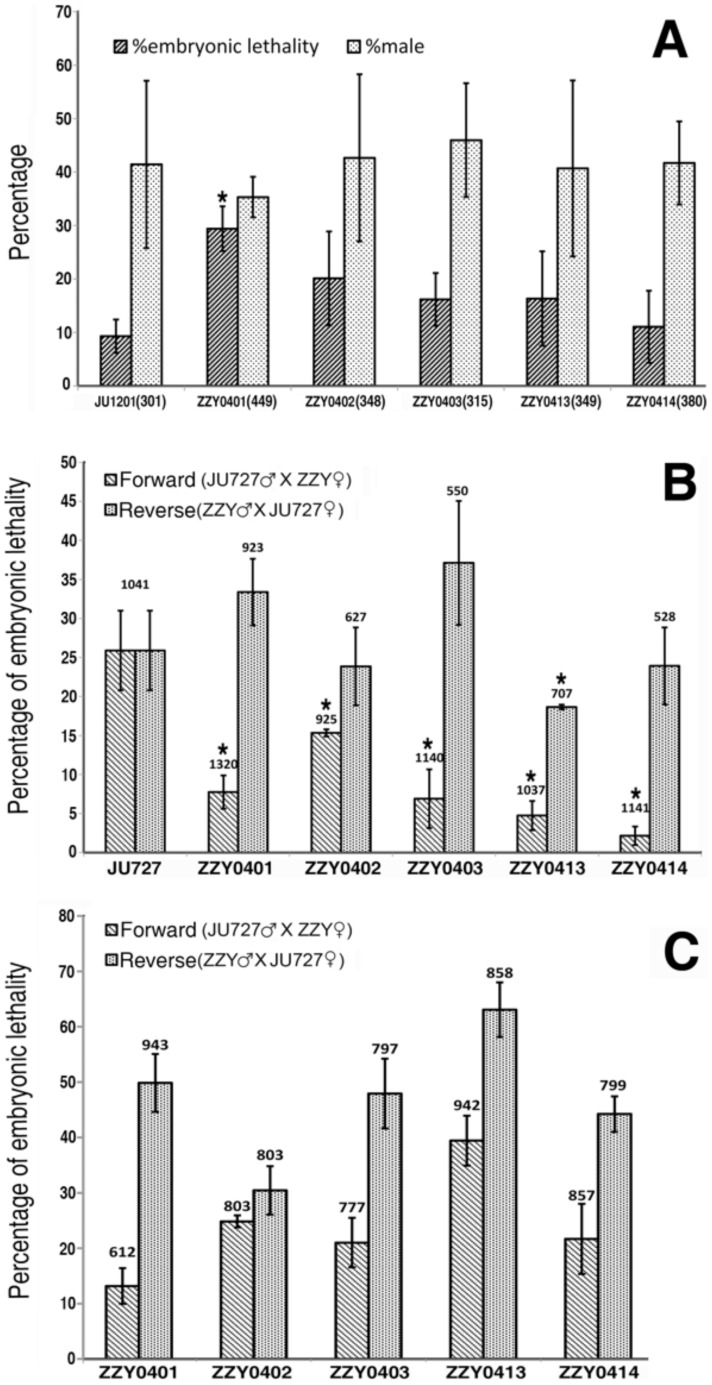
We re-isolated in China a relative of the nematode model Caenorhabditis elegans that was previously referred to informally as C. sp. 5. In spite of its importance for comparative biology, C. sp. 5 has remained morphologically uncharacterized. Therefore, we now provide detailed description of morphology and anatomy, assigning the name of Caenorhabditis sinica sp. n. to this nematode that is found frequently in China. C. sinica sp. n. belongs to the Elegans group in the genus Caenorhabditis, being phylogenetically close to C. briggsae although differing in reproductive mode. The gonochoristic C. sinica sp. n. displays two significantly larger distal parts of uteri filled with sperms in the female/hermaphroditic gonad than does the androdioecious C. briggsae. The new species can be differentiated morphologically from all known Caenorhabditis species within the Elegans group by presenting a uniquely shaped, three-pointed hook structure on the male precloacal lip. The lateral field of C. sinica sp. n. is marked by three ridges that are flanked by two additional incisures, sometimes appearing as five ridges in total. This study ends the prolonged period of the 'undescribed' anonymity for C. sinica sp. n. since its discovery and use in comparative biological research. Significant and crossing-direction dependent hybrid incompatibilities in F1 and F2 crossing progeny make C. sinica sp. n. an excellent model for studies of population and speciation genetics. The abundance of nematode species lacking detailed taxonomic characterization deserves renewed attention to address the species description gap for this important yet morphologically 'difficult' group of animals.
Conflict of interest statement
Figures

Similar articles
-
A phylogeny and molecular barcodes for Caenorhabditis, with numerous new species from rotting fruits.BMC Evol Biol. 2011 Nov 21;11:339. doi: 10.1186/1471-2148-11-339. BMC Evol Biol. 2011. PMID: 22103856 Free PMC article.
-
Genetic variation for postzygotic reproductive isolation between Caenorhabditis briggsae and Caenorhabditis sp. 9.Evolution. 2012 Apr;66(4):1180-95. doi: 10.1111/j.1558-5646.2011.01514.x. Epub 2011 Dec 12. Evolution. 2012. PMID: 22486697
-
Insights into species divergence and the evolution of hermaphroditism from fertile interspecies hybrids of Caenorhabditis nematodes.Genetics. 2010 Nov;186(3):997-1012. doi: 10.1534/genetics.110.120550. Epub 2010 Sep 7. Genetics. 2010. PMID: 20823339 Free PMC article.
-
The draft genome sequence of the nematode Caenorhabditis briggsae, a companion to C. elegans.Genome Biol. 2003;4(12):238. doi: 10.1186/gb-2003-4-12-238. Epub 2003 Nov 18. Genome Biol. 2003. PMID: 14659008 Free PMC article. Review.
-
Genomics and biology of the nematode Caenorhabditis briggsae.WormBook. 2007 May 3:1-16. doi: 10.1895/wormbook.1.136.1. WormBook. 2007. PMID: 18050493 Free PMC article. Review.
Cited by
-
Mutagenesis of GATA motifs controlling the endoderm regulator elt-2 reveals distinct dominant and secondary cis-regulatory elements.Dev Biol. 2016 Apr 1;412(1):160-170. doi: 10.1016/j.ydbio.2016.02.013. Epub 2016 Feb 16. Dev Biol. 2016. PMID: 26896592 Free PMC article.
-
Deep sampling of Hawaiian Caenorhabditis elegans reveals high genetic diversity and admixture with global populations.Elife. 2019 Dec 3;8:e50465. doi: 10.7554/eLife.50465. Elife. 2019. PMID: 31793880 Free PMC article.
-
MIPhy: identify and quantify rapidly evolving members of large gene families.PeerJ. 2018 May 29;6:e4873. doi: 10.7717/peerj.4873. eCollection 2018. PeerJ. 2018. PMID: 29868279 Free PMC article.
-
Specific Interactions Between Autosome and X Chromosomes Cause Hybrid Male Sterility in Caenorhabditis Species.Genetics. 2019 Jul;212(3):801-813. doi: 10.1534/genetics.119.302202. Epub 2019 May 7. Genetics. 2019. PMID: 31064822 Free PMC article.
-
Five new Caenorhabditis species from Indonesia provide exceptions to Haldane's rule and partial fertility of interspecific hybrids.G3 (Bethesda). 2025 Aug 6;15(8):jkaf134. doi: 10.1093/g3journal/jkaf134. G3 (Bethesda). 2025. PMID: 40542721 Free PMC article.
References
-
- Kiontke K, Sudhaus W (2006) Ecology of Caenorhabditis species. In: The C. elegans Research Community, editor. Wormbook. (January 09, 2006) ed. doi/10.1895/wormbook.1.37.1: http://www.wormbook.org. - PMC - PubMed
-
- Félix M-A (2007) Cryptic quantitative evolution of the vulva intercellular signaling network in Caenorhabditis . Current Biology 17: 103–114. - PubMed
-
- Kiontke K, Barriere A, Kolotuev I, Podbilewicz B, Sommer R, et al. (2007) Trends, stasis, and drift in the evolution of nematode vulva development. Current Biology 17: 1925–1937. - PubMed
-
- Cutter AD (2008) Divergence times in Caenorhabditis and Drosophila inferred from direct estimates of the neutral mutation rate. Mol Biol Evol 25: 778–786. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
