

Human Fungal Pathogens of Mucorales and Entomophthorales
- PMID: 25377138
- PMCID: PMC4382724
- DOI: 10.1101/cshperspect.a019562
Human Fungal Pathogens of Mucorales and Entomophthorales
Abstract
In recent years, we have seen an increase in the number of immunocompromised cohorts as a result of infections and/or medical conditions, which has resulted in an increased incidence of fungal infections. Although rare, the incidence of infections caused by fungi belonging to basal fungal lineages is also continuously increasing. Basal fungal lineages diverged at an early point during the evolution of the fungal lineage, in which, in a simplified four-phylum fungal kingdom, Zygomycota and Chytridiomycota belong to the basal fungi, distinguishing them from Ascomycota and Basidiomycota. Currently there are no known human infections caused by fungi in Chytridiomycota; only Zygomycotan fungi are known to infect humans. Hence, infections caused by zygomycetes have been called zygomycosis, and the term "zygomycosis" is often used as a synonym for "mucormycosis." In the four-phylum fungal kingdom system, Zygomycota is classified mainly based on morphology, including the ability to form coenocytic (aseptated) hyphae and zygospores (sexual spores). In the Zygomycota, there are 10 known orders, two of which, the Mucorales and Entomophthorales, contain species that can infect humans, and the infection has historically been known as zygomycosis. However, recent multilocus sequence typing analyses (the fungal tree of life [AFTOL] project) revealed that the Zygomycota forms not a monophyletic clade but instead a polyphyletic clade, whereas Ascomycota and Basidiomycota are monophyletic. Thus, the term "zygomycosis" needed to be further specified, resulting in the terms "mucormycosis" and "entomophthoramycosis." This review covers these two different types of fungal infections.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
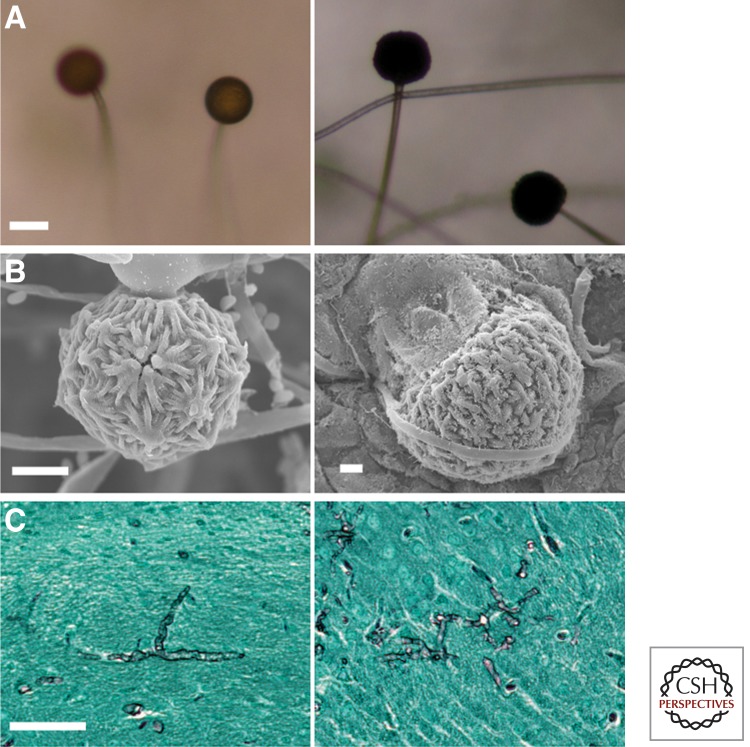
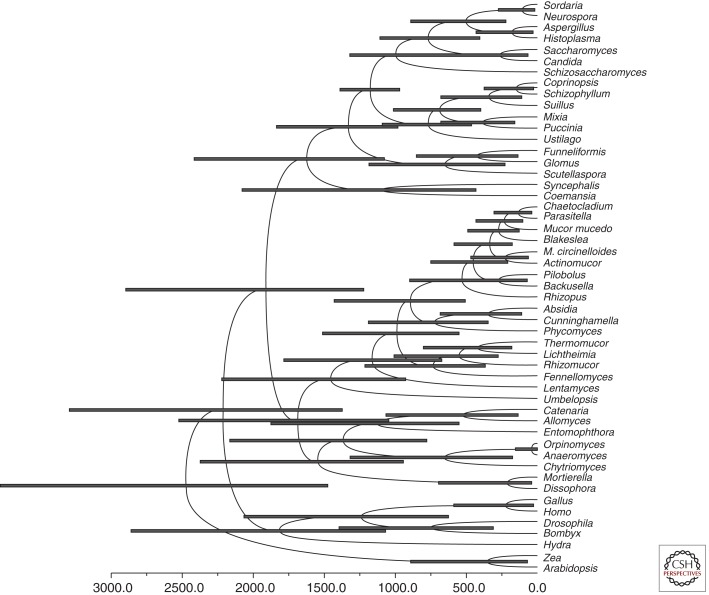
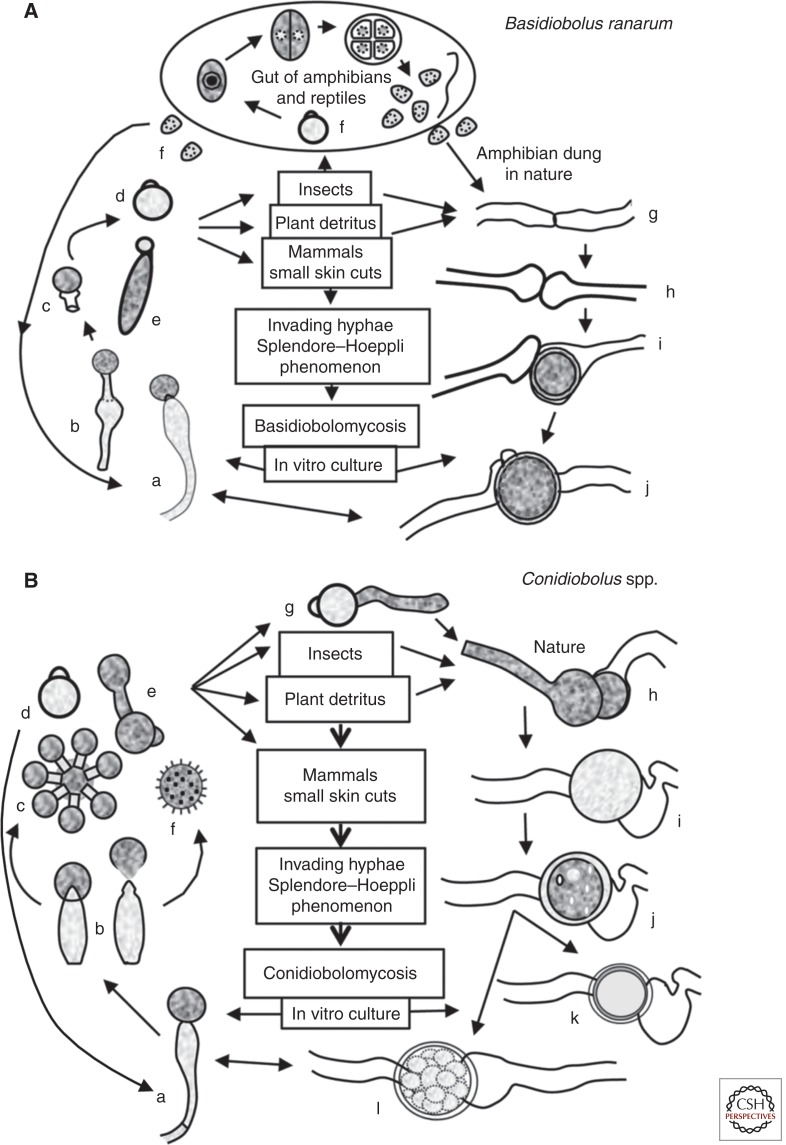
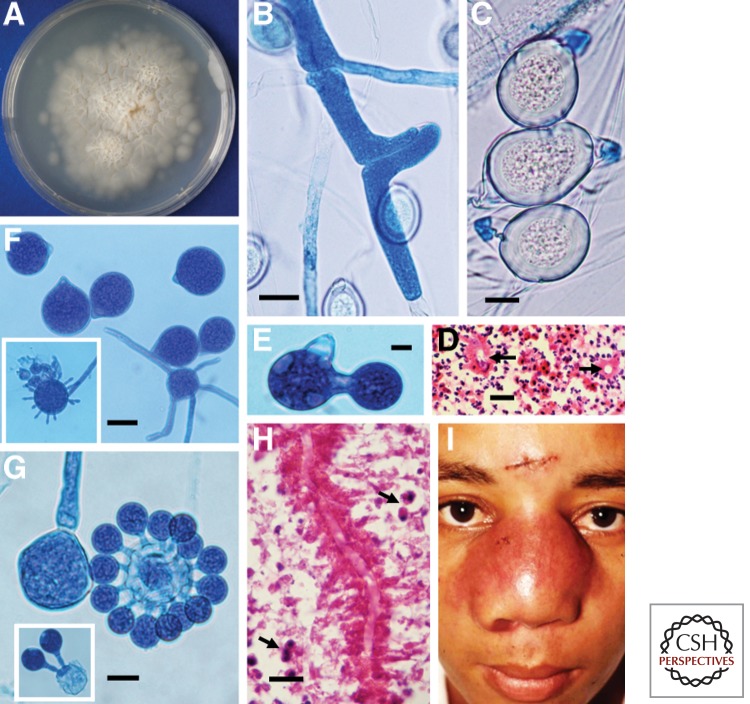

References
-
- Abe F, Inaba H, Katoh T, Hotchi M 1990. Effects of iron and desferrioxamine on Rhizopus infection. Mycopathologia 110: 87–91. - PubMed
-
- Abe A, Oda Y, Asano K, Sone T 2007. Rhizopus delemar is the proper name for Rhizopus oryzae fumaric-malic acid producers. Mycologia 99: 714–722. - PubMed
-
- Abzug MJ, Walsh TJ 2004. Interferon-γ and colony-stimulating factors as adjuvant therapy for refractory fungal infections in children. Pediatr Infect Dis J 23: 769–773. - PubMed
-
- Al-Hajjar S, Perfect J, Hashem F, Tufenkeji H, Kayes S 1996. Orbitofacial conidiobolomycosis in a child. Pediatr Infect Dis J 15: 1130–1132. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources