

Caspr and caspr2 are required for both radial and longitudinal organization of myelinated axons
- PMID: 25378149
- PMCID: PMC4220019
- DOI: 10.1523/JNEUROSCI.3369-14.2014
Caspr and caspr2 are required for both radial and longitudinal organization of myelinated axons
Abstract
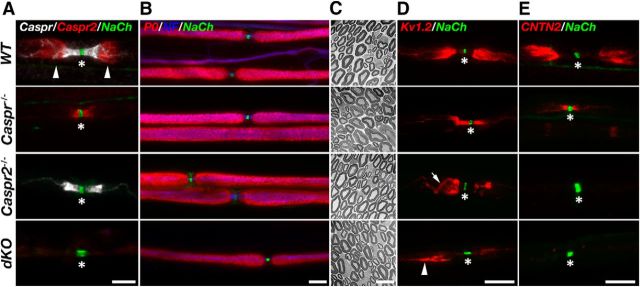
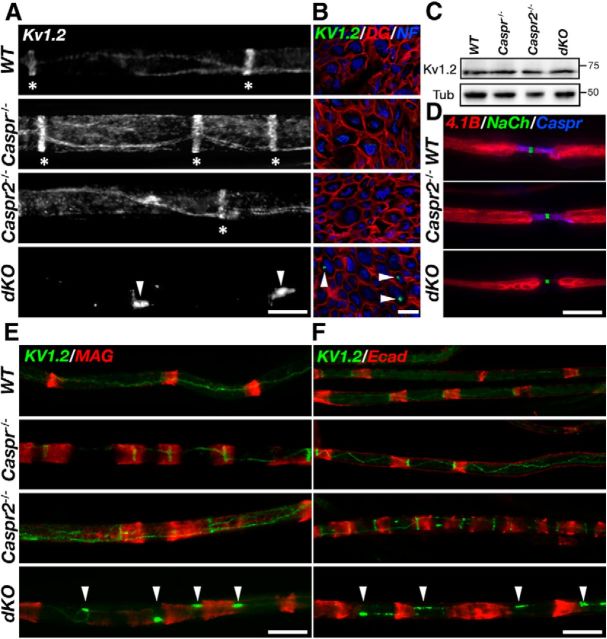
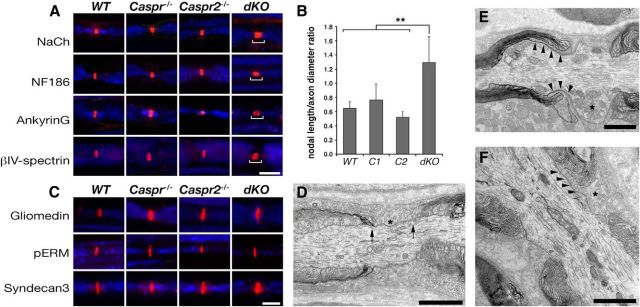
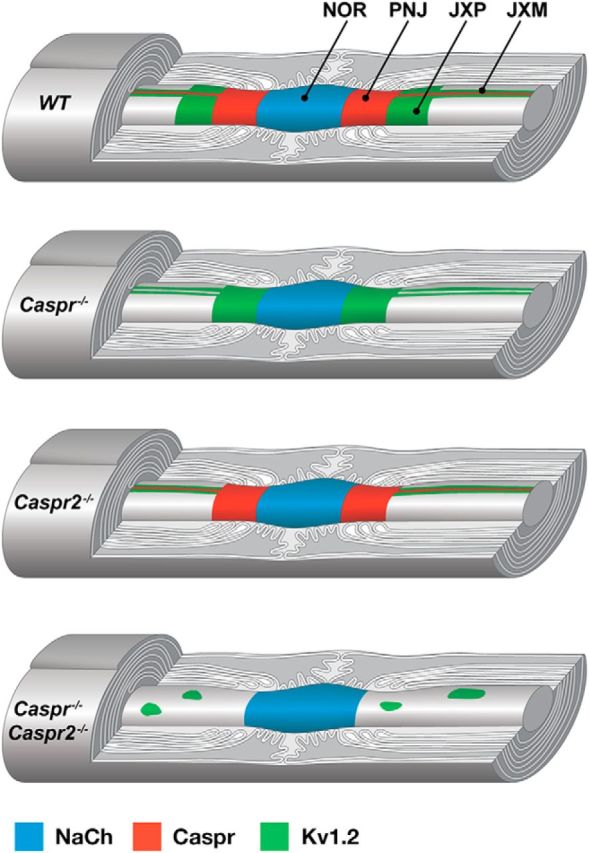
In myelinated peripheral axons, Kv1 potassium channels are clustered at the juxtaparanodal region and at an internodal line located along the mesaxon and below the Schmidt-Lanterman incisures. This polarized distribution is controlled by Schwann cells and requires specific cell adhesion molecules (CAMs). The accumulation of Kv1 channels at the juxtaparanodal region depends on the presence of Caspr2 at this site, as well as on the presence of Caspr at the adjacent paranodal junction. However, the localization of these channels along the mesaxonal internodal line still persists in the absence of each one of these CAMs. By generating mice lacking both Caspr and Caspr2 (caspr(-/-)/caspr2(-/-)), we now reveal compensatory functions of the two proteins in the organization of the axolemma. Although Kv1 channels are clustered along the inner mesaxon and in a circumferential ring below the incisures in the single mutants, in sciatic nerves of caspr(-/-)/caspr2(-/-) mice, these channels formed large aggregates that were dispersed along the axolemma, demonstrating that internodal localization of Kv1 channels requires either Caspr or Caspr2. Furthermore, deletion of both Caspr and Caspr2 also resulted in widening of the nodes of Ranvier, suggesting that Caspr2 (which is present at paranodes in the absence of Caspr) can partially compensate for the barrier function of Caspr at this site even without the formation of a distinct paranodal junction. Our results indicate that Caspr and Caspr2 are required for the organization of the axolemma both radially, manifested as the mesaxonal line, and longitudinally, demarcated by the nodal domains.
Keywords: Caspr; adhesion molecule; axon–glia interaction; myelin; node of Ranvier; peripheral nerve.
Copyright © 2014 the authors 0270-6474/14/3414820-07$15.00/0.
Figures
References
-
- Amor V, Feinberg K, Eshed-Eisenbach Y, Vainshtein A, Frechter S, Grumet M, Rosenbluth J, Peles E. Long-term maintenance of Na+ channels at nodes of Ranvier depends on glial contact mediated by gliomedin and NrCAM. J Neurosci. 2014;34:5089–5098. doi: 10.1523/JNEUROSCI.4752-13.2014. - DOI - PMC - PubMed
-
- Bhat MA, Rios JC, Lu Y, Garcia-Fresco GP, Ching W, St Martin M, Li J, Einheber S, Chesler M, Rosenbluth J, Salzer JL, Bellen HJ. Axon-glia interactions and the domain organization of myelinated axons requires neurexin IV/Caspr/Paranodin. Neuron. 2001;30:369–383. doi: 10.1016/S0896-6273(01)00294-X. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases