

The influenza virus neuraminidase protein transmembrane and head domains have coevolved
- PMID: 25378494
- PMCID: PMC4300628
- DOI: 10.1128/JVI.02005-14
The influenza virus neuraminidase protein transmembrane and head domains have coevolved
Abstract
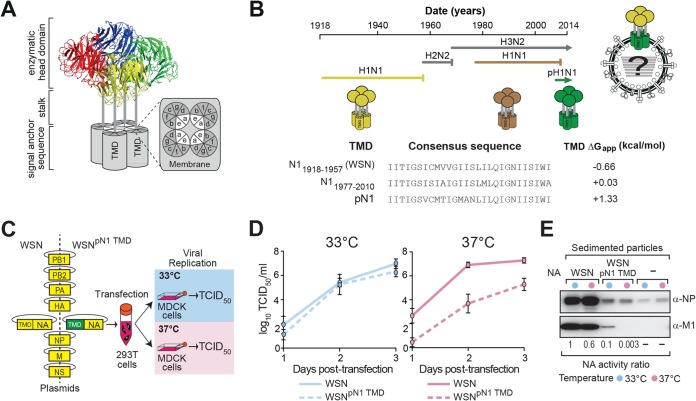
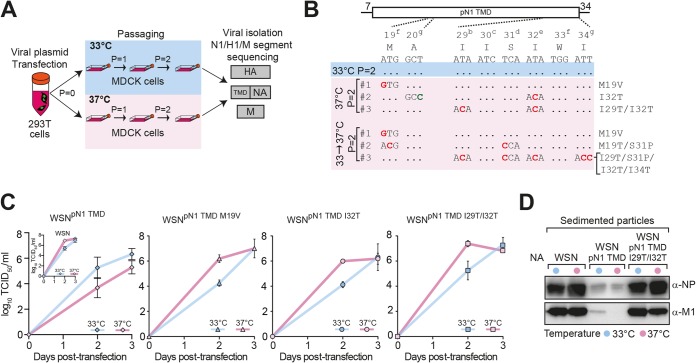
Transmembrane domains (TMDs) from single-spanning membrane proteins are commonly viewed as membrane anchors for functional domains. Influenza virus neuraminidase (NA) exemplifies this concept, as it retains enzymatic function upon proteolytic release from the membrane. However, the subtype 1 NA TMDs have become increasingly more polar in human strains since 1918, which suggests that selection pressure exists on this domain. Here, we investigated the N1 TMD-head domain relationship by exchanging a prototypical "old" TMD (1933) with a "recent" (2009), more polar TMD and an engineered hydrophobic TMD. Each exchange altered the TMD association, decreased the NA folding efficiency, and significantly reduced viral budding and replication at 37°C compared to at 33°C, at which NA folds more efficiently. Passaging the chimera viruses at 37°C restored the NA folding efficiency, viral budding, and infectivity by selecting for NA TMD mutations that correspond with their polar or hydrophobic assembly properties. These results demonstrate that single-spanning membrane protein TMDs can influence distal domain folding, as well as membrane-related processes, and suggest the NA TMD in H1N1 viruses has become more polar to maintain compatibility with the evolving enzymatic head domain.
Importance: The neuraminidase (NA) protein from influenza A viruses (IAVs) functions to promote viral release and is one of the major surface antigens. The receptor-destroying activity in NA resides in the distal head domain that is linked to the viral membrane by an N-terminal hydrophobic transmembrane domain (TMD). Over the last century, the subtype 1 NA TMDs (N1) in human H1N1 viruses have become increasingly more polar, and the head domains have changed to alter their antigenicity. Here, we provide the first evidence that an "old" N1 head domain from 1933 is incompatible with a "recent" (2009), more polar N1 TMD sequence and that, during viral replication, the head domain drives the selection of TMD mutations. These mutations modify the intrinsic TMD assembly to restore the head domain folding compatibility and the resultant budding deficiency. This likely explains why the N1 TMDs have become more polar and suggests the N1 TMD and head domain have coevolved.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
