

Principles of ER cotranslational translocation revealed by proximity-specific ribosome profiling
- PMID: 25378630
- PMCID: PMC4285348
- DOI: 10.1126/science.1257521
Principles of ER cotranslational translocation revealed by proximity-specific ribosome profiling
Abstract
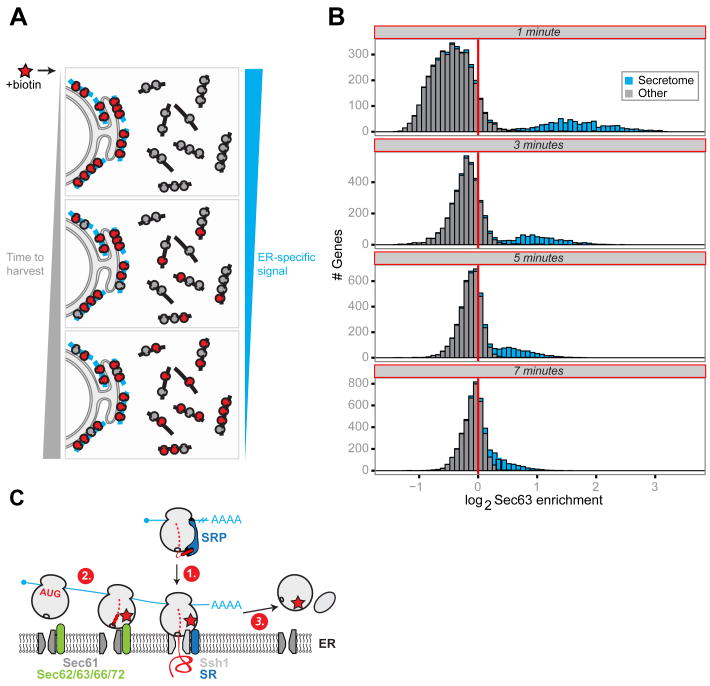
Localized protein synthesis is a fundamental mechanism for creating distinct subcellular environments. Here we developed a generalizable proximity-specific ribosome profiling strategy that enables global analysis of translation in defined subcellular locations. We applied this approach to the endoplasmic reticulum (ER) in yeast and mammals. We observed the large majority of secretory proteins to be cotranslationally translocated, including substrates capable of posttranslational insertion in vitro. Distinct translocon complexes engaged nascent chains at different points during synthesis. Whereas most proteins engaged the ER immediately after or even before signal sequence (SS) emergence, a class of Sec66-dependent proteins entered with a looped SS conformation. Finally, we observed rapid ribosome exchange into the cytosol after translation termination. These data provide insights into how distinct translocation mechanisms act in concert to promote efficient cotranslational recruitment.
Copyright © 2014, American Association for the Advancement of Science.
Figures
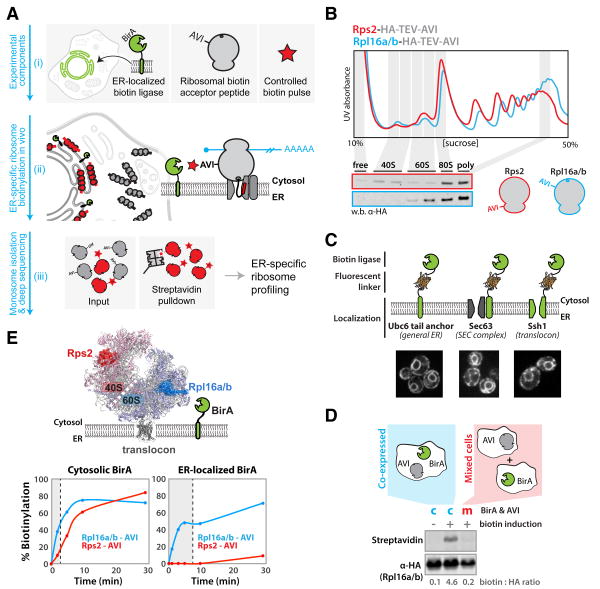
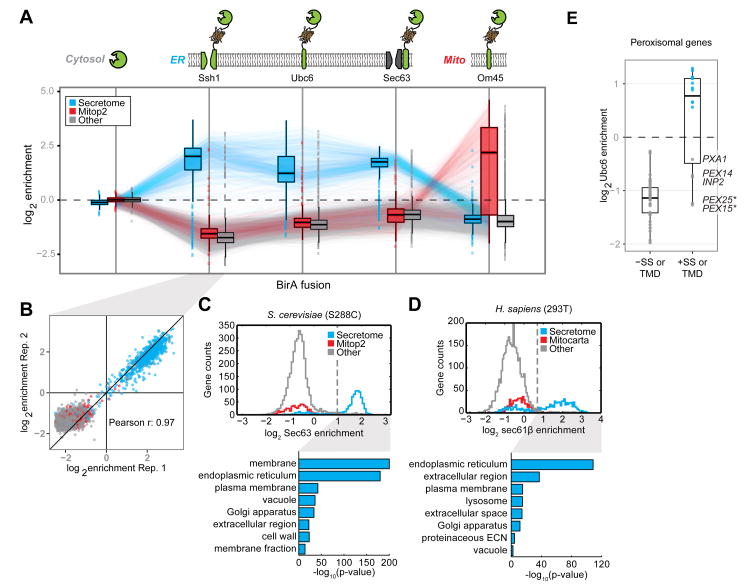
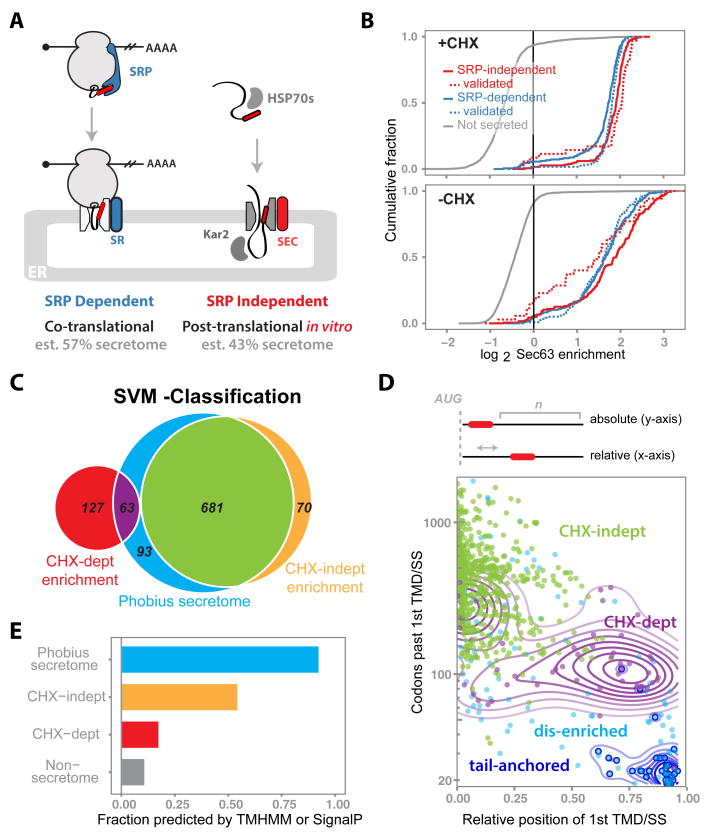
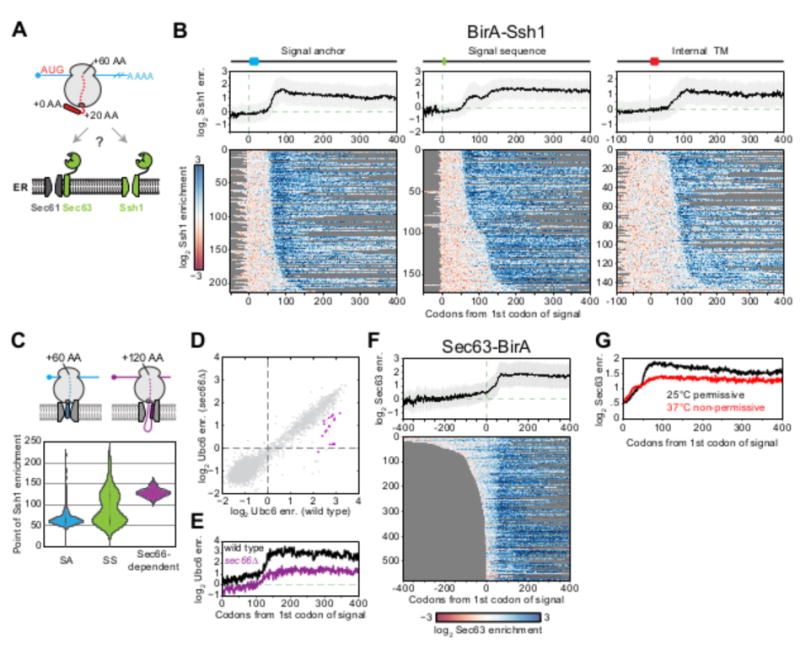
Comment in
-
Cell Biology. Local synthesis and disposal.Science. 2014 Nov 7;346(6210):701-2. doi: 10.1126/science.1261602. Science. 2014. PMID: 25378608 No abstract available.
-
A global look at local translation.Nat Methods. 2015 Jan;12(1):17. doi: 10.1038/nmeth.3247. Nat Methods. 2015. PMID: 25699318 No abstract available.
-
LOCAL TRANSLATION. Comment on "Principles of ER cotranslational translocation revealed by proximity-specific ribosome profiling".Science. 2015 Jun 12;348(6240):1217. doi: 10.1126/science.aaa7257. Epub 2015 Jun 11. Science. 2015. PMID: 26068841 Free PMC article.
-
LOCAL TRANSLATION. Response to Comment on "Principles of ER cotranslational translocation revealed by proximity-specific ribosome profiling".Science. 2015 Jun 12;348(6240):1217. doi: 10.1126/science.aaa8299. Epub 2015 Jun 11. Science. 2015. PMID: 26068842
References
-
- St Johnston D. Moving messages: the intracellular localization of mRNAs. Nat Rev Mol Cell Biol. 2005;6:363–375. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
