

Relative roles of ABCG5/ABCG8 in liver and intestine
- PMID: 25378657
- PMCID: PMC4306686
- DOI: 10.1194/jlr.M054544
Relative roles of ABCG5/ABCG8 in liver and intestine
Abstract
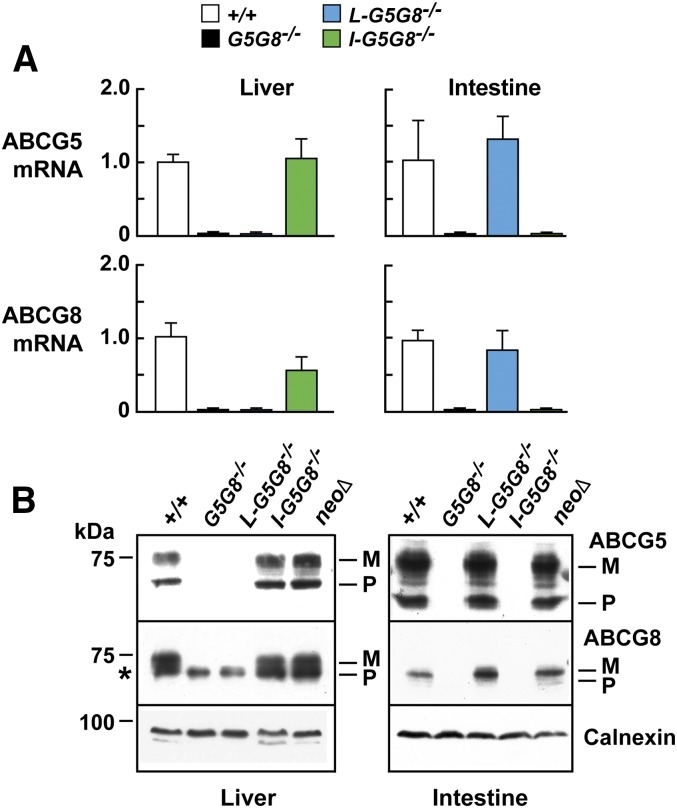
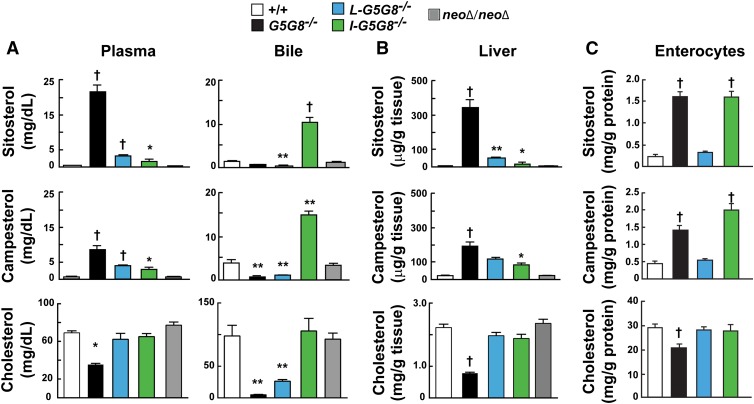
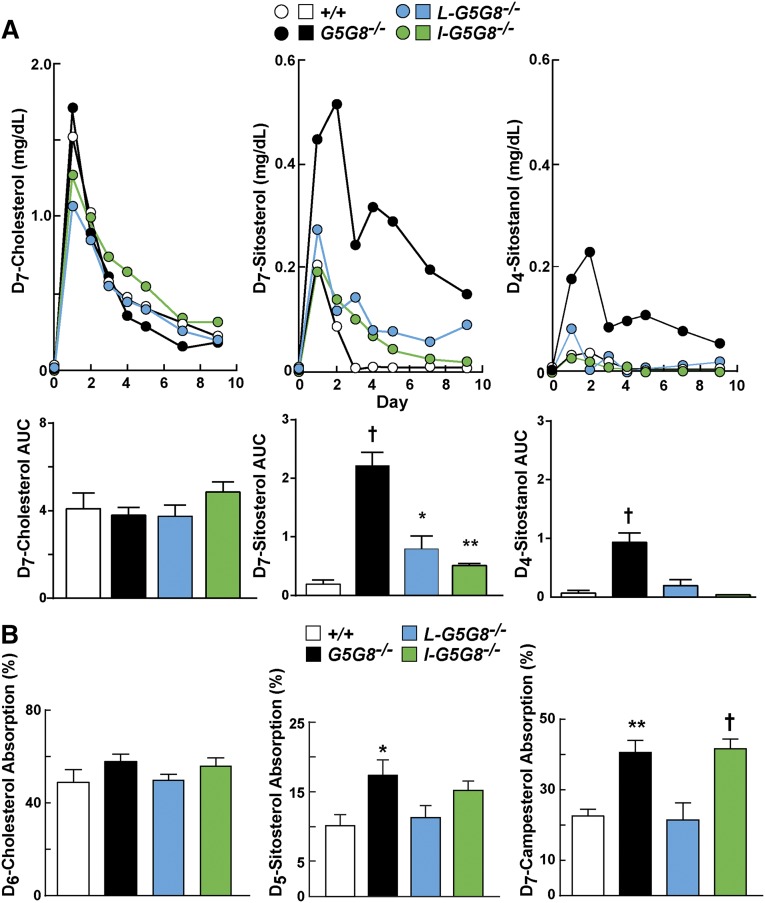
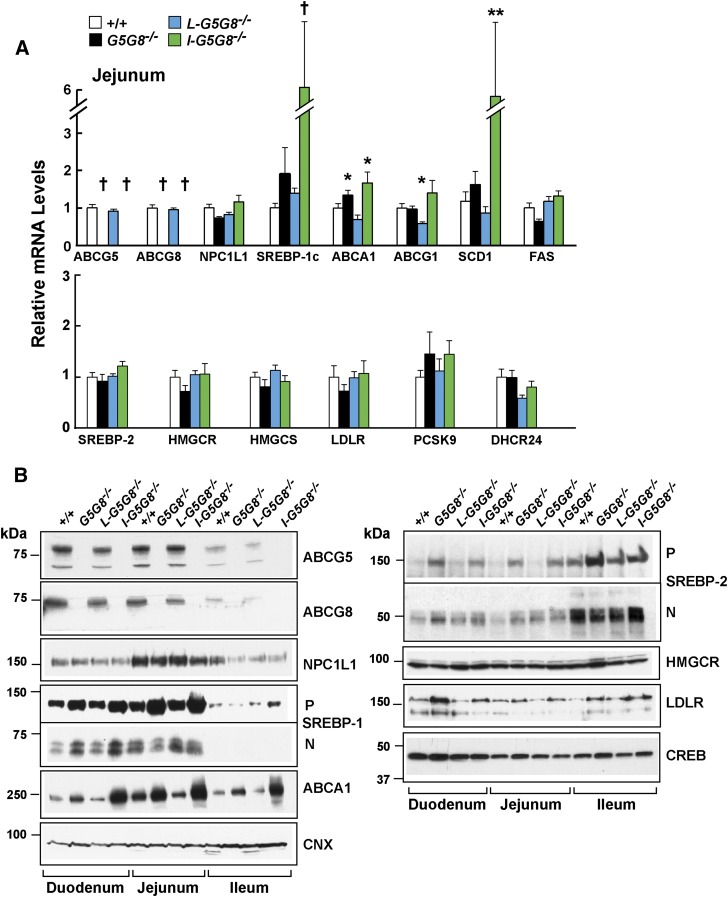
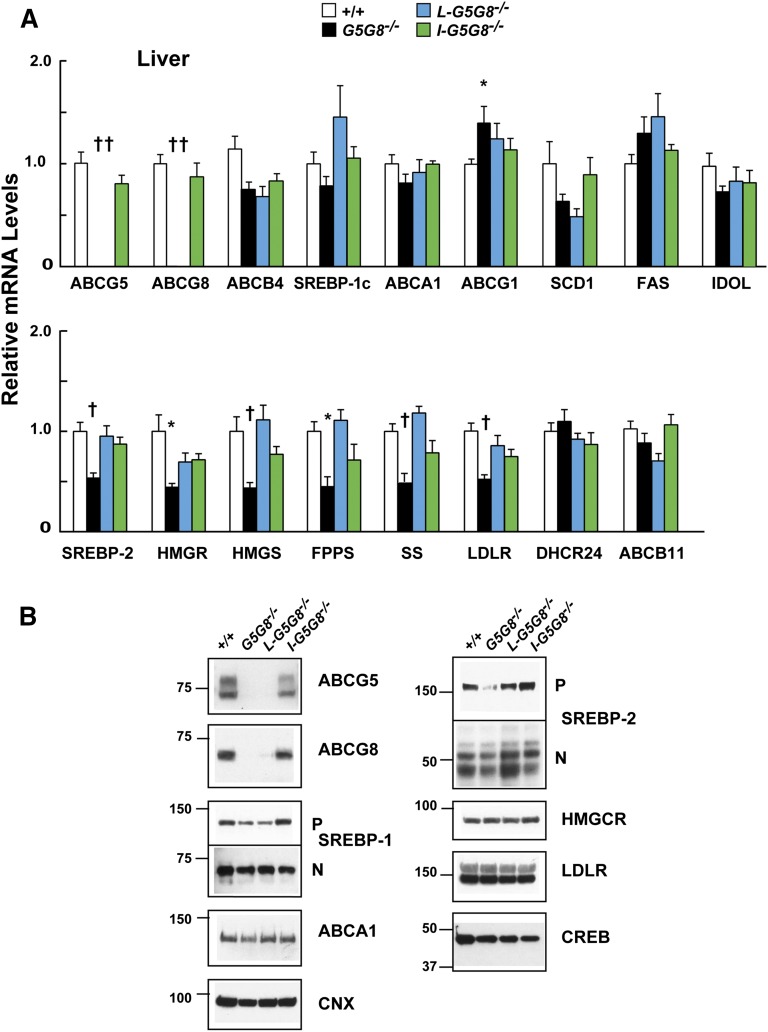
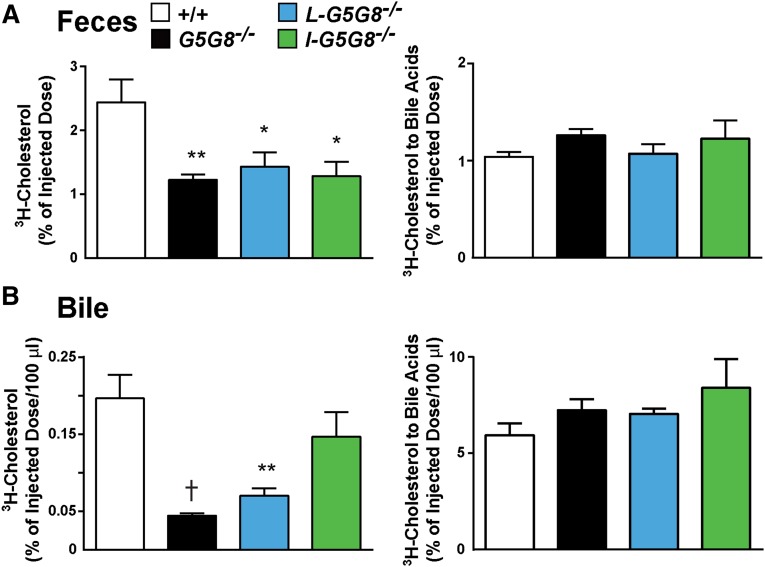
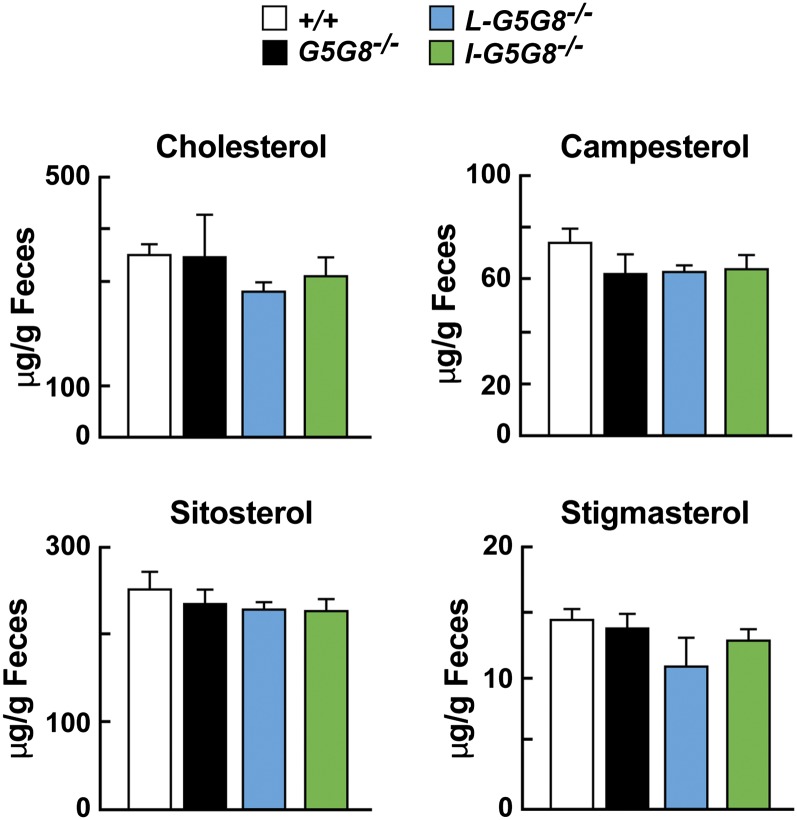
ABCG5 (G5) and ABCG8 (G8) form a sterol transporter that acts in liver and intestine to prevent accumulation of dietary sterols. Mutations in either G5 or G8 cause sitosterolemia, a recessive disorder characterized by sterol accumulation and premature coronary atherosclerosis. Hepatic G5G8 mediates cholesterol excretion into bile, but the function and relative importance of intestinal G5G8 has not been defined. To determine the role of intestinal G5G8, we developed liver-specific (L-G5G8(-/-)), intestine-specific (I-G5G8(-/-)), and total (G5G8(-/-)) KO mice. Tissue levels of sitosterol, the most abundant plant sterol, were >90-fold higher in G5G8(-/-) mice than in WT animals. Expression of G5G8 only in intestine or only in liver decreased tissue sterol levels by 90% when compared with G5G8(-/-) animals. Biliary sterol secretion was reduced in L-G5G8(-/-) and G5G8(-/-) mice, but not in I-G5G8(-/-) mice. Conversely, absorption of plant sterols was increased in I-G5G8(-/-) and G5G8(-/-) mice, but not in L-G5G8(-/-) mice. Reverse cholesterol transport, as assessed from the fraction of intravenously administered (3)H-cholesterol that appeared in feces, was reduced in G5G8(-/-), I-G5G8(-/-), and L-G5G8(-/-) mice. Thus, G5G8 expression in both the liver and intestine protects animals from sterol accumulation, and intestinal G5G8 contributes to extrahepatic cholesterol efflux in mice.
Keywords: bile; cholesterol/absorption; sterols; transport.
Copyright © 2015 by the American Society for Biochemistry and Molecular Biology, Inc.
Figures
Comment in
-
Hepatic or intestinal ABCG5 and ABCG8 are sufficient to block the development of sitosterolemia.J Lipid Res. 2015 Feb;56(2):201-2. doi: 10.1194/jlr.C056945. Epub 2014 Dec 19. J Lipid Res. 2015. PMID: 25527605 Free PMC article. No abstract available.
References
-
- Miettinen T. A. 1980. Phytosterolaemia, xanthomatosis and premature atherosclerotic arterial disease: a case with high plant sterol absorption, impaired sterol elimination and low cholesterol synthesis. Eur. J. Clin. Invest. 10: 27–35. - PubMed
-
- Berge K. E., Tian H., Graf G. A., Yu L., Grishin N. V., Schultz J., Kwiterovich P., Shan B., Barnes R., Hobbs H. H. 2000. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science. 290: 1771–1775. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
