

Insights into the specificity of lysine acetyltransferases
- PMID: 25381442
- PMCID: PMC4276886
- DOI: 10.1074/jbc.M114.613901
Insights into the specificity of lysine acetyltransferases
Abstract
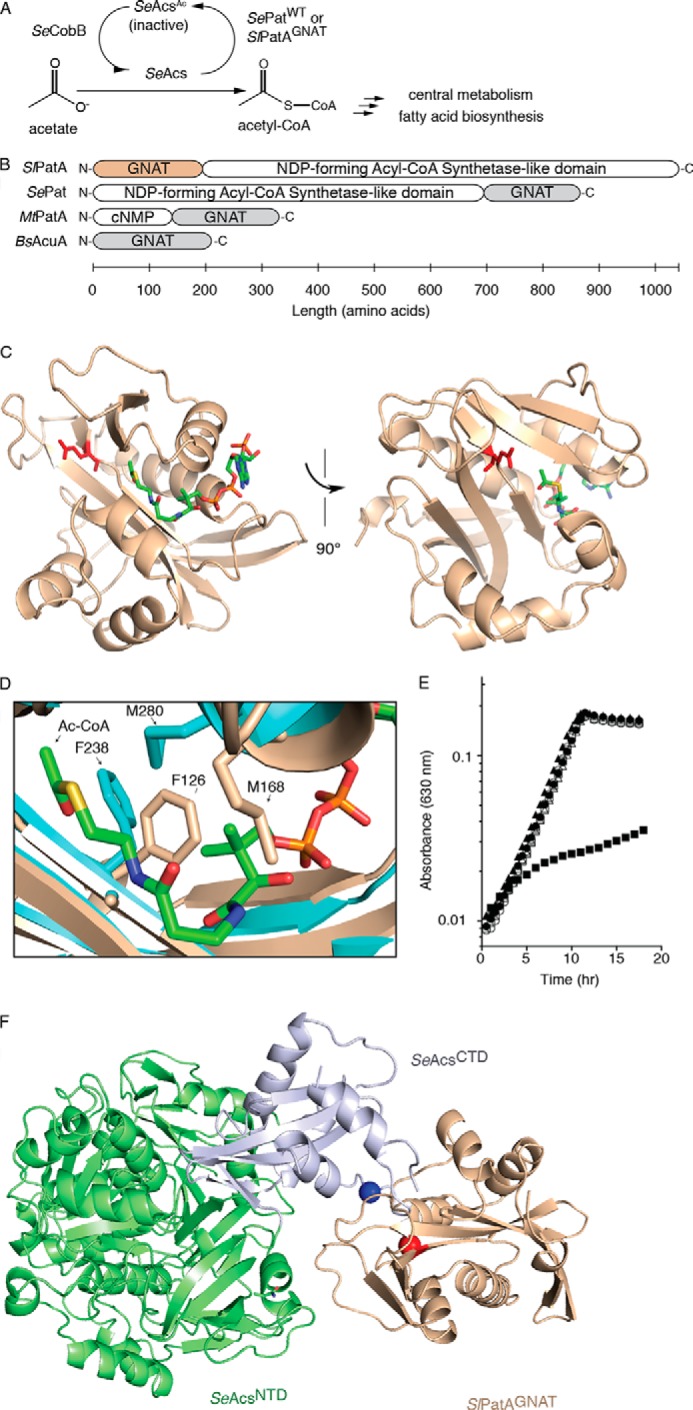
Reversible lysine acetylation by protein acetyltransferases is a conserved regulatory mechanism that controls diverse cellular pathways. Gcn5-related N-acetyltransferases (GNATs), named after their founding member, are found in all domains of life. GNATs are known for their role as histone acetyltransferases, but non-histone bacterial protein acetytransferases have been identified. Only structures of GNAT complexes with short histone peptide substrates are available in databases. Given the biological importance of this modification and the abundance of lysine in polypeptides, how specificity is attained for larger protein substrates is central to understanding acetyl-lysine-regulated networks. Here we report the structure of a GNAT in complex with a globular protein substrate solved to 1.9 Å. GNAT binds the protein substrate with extensive surface interactions distinct from those reported for GNAT-peptide complexes. Our data reveal determinants needed for the recognition of a protein substrate and provide insight into the specificity of GNATs.
Keywords: Acetyl Coenzyme A (Acetyl-CoA); Bacterial Metabolism; Enzyme Inactivation; Histone Acetylase; Post-translational Modification (PTM); Substrate Specificity.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures






References
-
- Kouzarides T. (2007) Chromatin modifications and their function. Cell 128, 693–705 - PubMed
-
- Glozak M. A., Sengupta N., Zhang X., Seto E. (2005) Acetylation and deacetylation of non-histone proteins. Gene 363, 15–23 - PubMed
-
- Hubbert C., Guardiola A., Shao R., Kawaguchi Y., Ito A., Nixon A., Yoshida M., Wang X. F., Yao T. P. (2002) HDAC6 is a microtubule-associated deacetylase. Nature 417, 455–458 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
