

Germinal center reentries of BCL2-overexpressing B cells drive follicular lymphoma progression
- PMID: 25384217
- PMCID: PMC4348942
- DOI: 10.1172/JCI72415
Germinal center reentries of BCL2-overexpressing B cells drive follicular lymphoma progression
Abstract
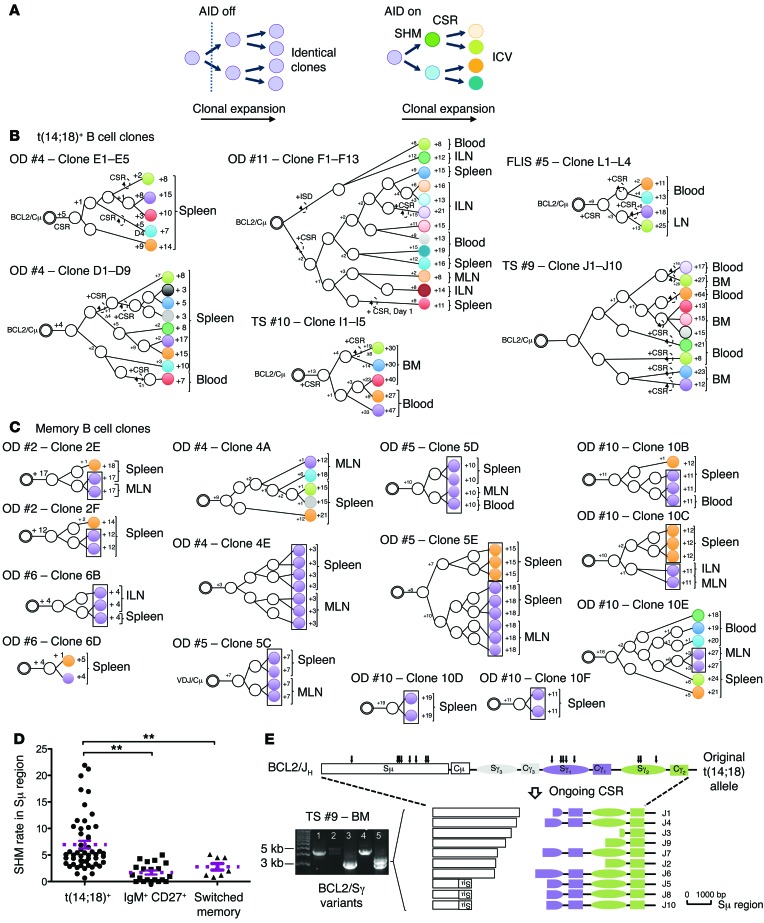
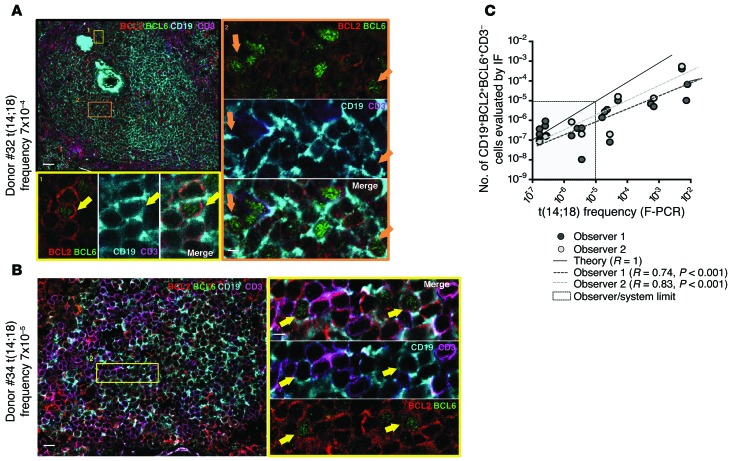
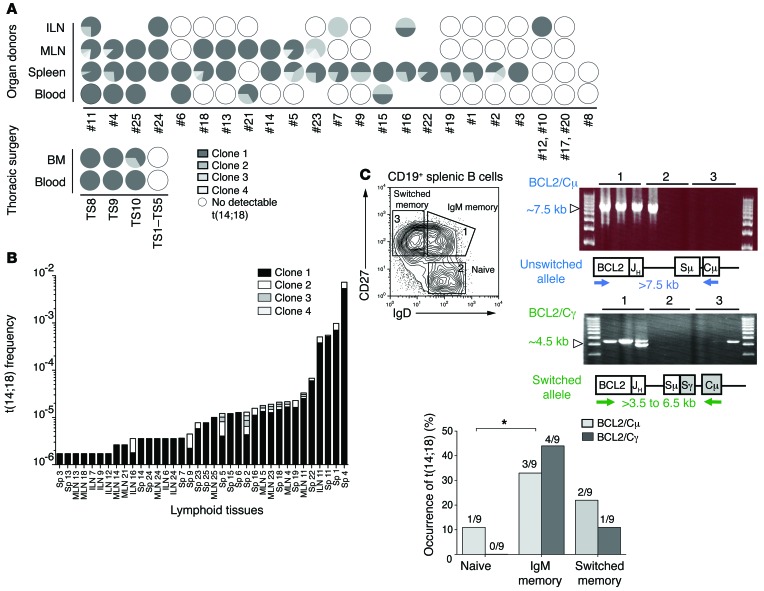
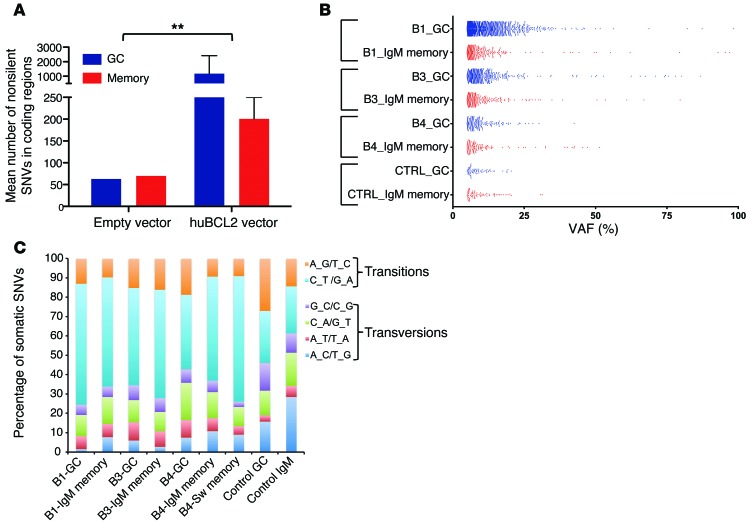
It has recently been demonstrated that memory B cells can reenter and reengage germinal center (GC) reactions, opening the possibility that multi-hit lymphomagenesis gradually occurs throughout life during successive immunological challenges. Here, we investigated this scenario in follicular lymphoma (FL), an indolent GC-derived malignancy. We developed a mouse model that recapitulates the FL hallmark t(14;18) translocation, which results in constitutive activation of antiapoptotic protein B cell lymphoma 2 (BCL2) in a subset of B cells, and applied a combination of molecular and immunofluorescence approaches to track normal and t(14;18)(+) memory B cells in human and BCL2-overexpressing B cells in murine lymphoid tissues. BCL2-overexpressing B cells required multiple GC transits before acquiring FL-associated developmental arrest and presenting as GC B cells with constitutive activation-induced cytidine deaminase (AID) mutator activity. Moreover, multiple reentries into the GC were necessary for the progression to advanced precursor stages of FL. Together, our results demonstrate that protracted subversion of immune dynamics contributes to early dissemination and progression of t(14;18)(+) precursors and shapes the systemic presentation of FL patients.
Figures

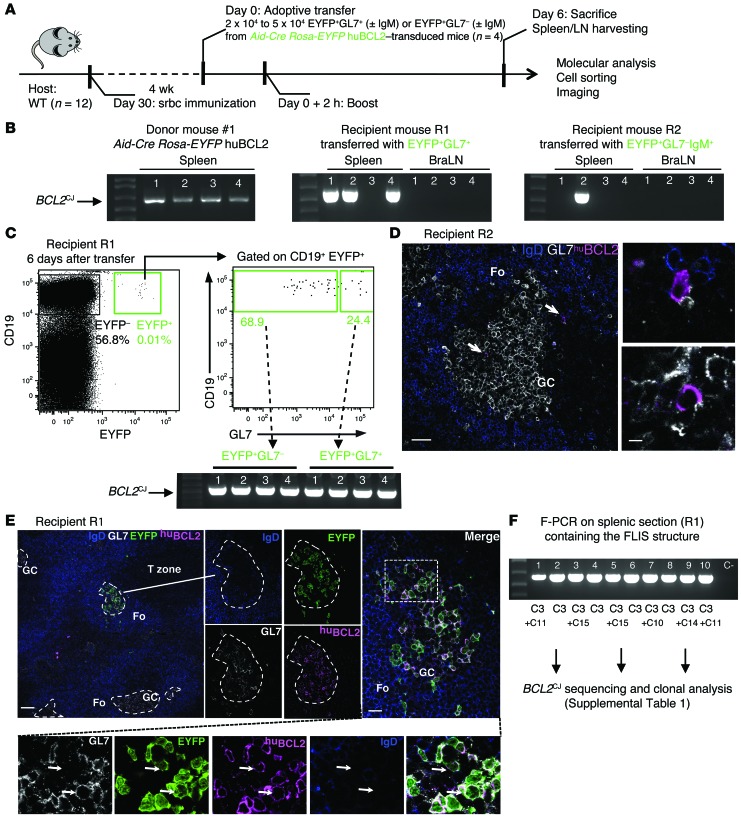
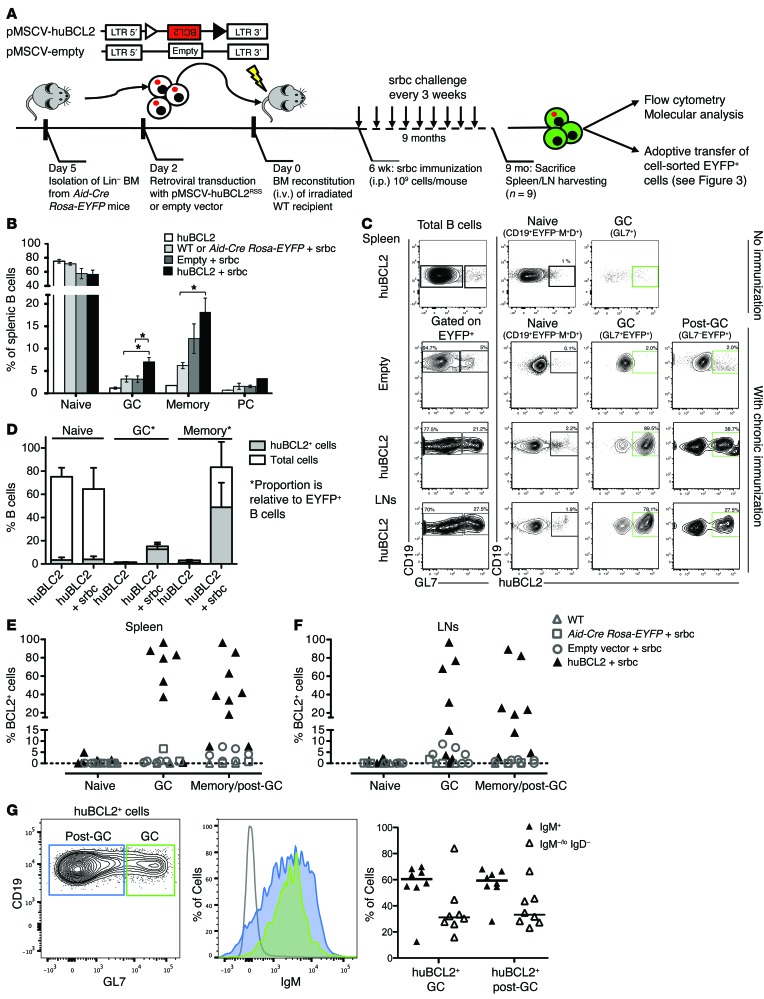
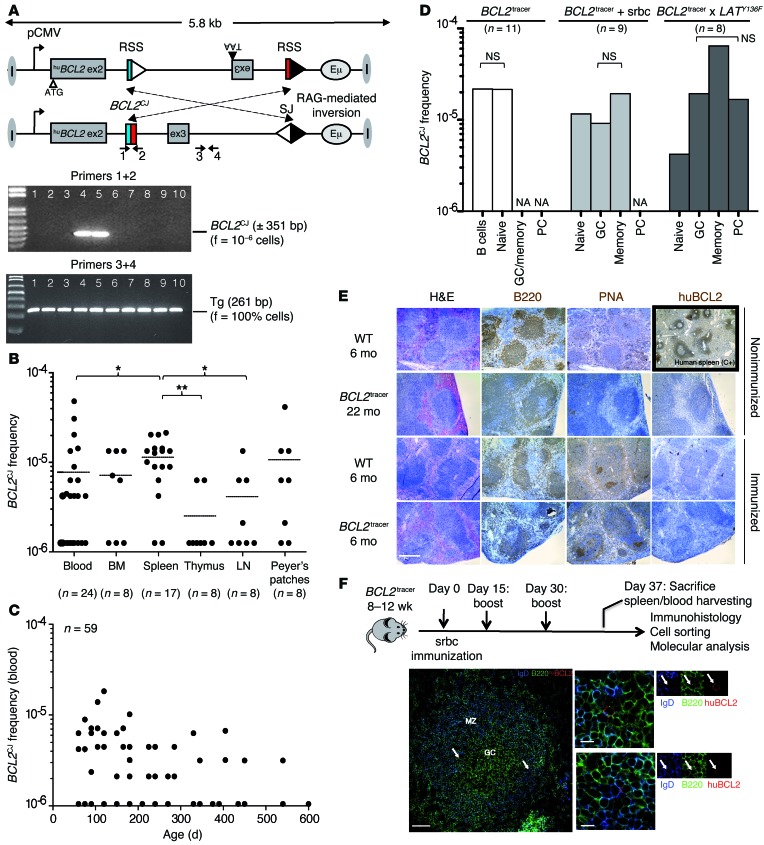
Comment in
-
Follicular lymphoma: too many reminders for a memory B cell.J Clin Invest. 2014 Dec;124(12):5095-8. doi: 10.1172/JCI79189. Epub 2014 Nov 10. J Clin Invest. 2014. PMID: 25384212 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
