

Thermally Dimorphic Human Fungal Pathogens--Polyphyletic Pathogens with a Convergent Pathogenicity Trait
- PMID: 25384771
- PMCID: PMC4526722
- DOI: 10.1101/cshperspect.a019794
Thermally Dimorphic Human Fungal Pathogens--Polyphyletic Pathogens with a Convergent Pathogenicity Trait
Abstract
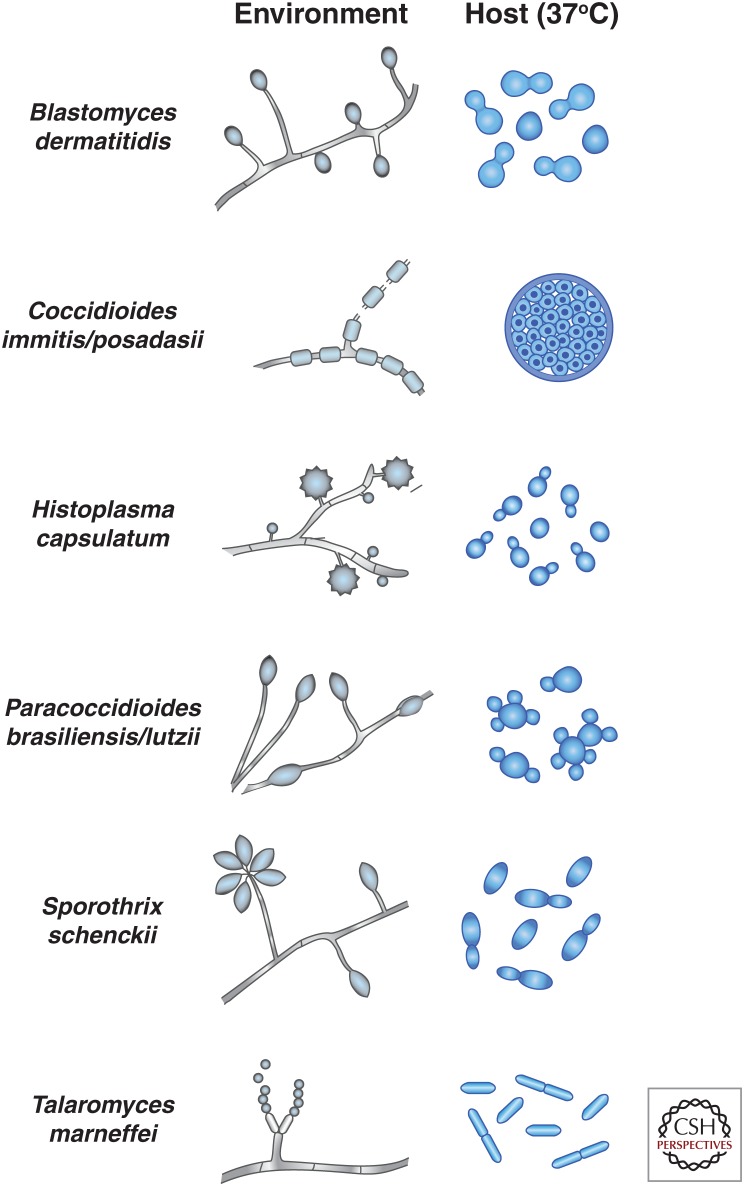
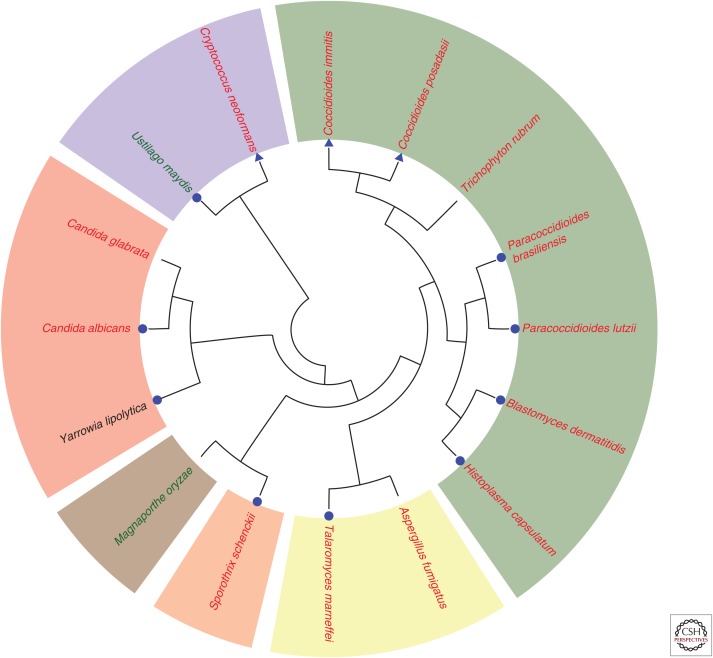
Fungi are adept at changing their cell shape and developmental program in response to signals in their surroundings. Here we focus on a group of evolutionarily related fungal pathogens of humans known as the thermally dimorphic fungi. These organisms grow in a hyphal form in the environment but shift their morphology drastically within a mammalian host. Temperature is one of the main host signals that initiates their conversion to the "host" form and is sufficient in the laboratory to trigger establishment of this host-adapted developmental program. Here we discuss the major human pathogens in this group, which are Blastomyces dermatiditis, Coccidioides immitis/posadasii, Histoplasma capsulatum, Paracoccidioides brasiliensis/lutzii, Sporothrix schenckii, and Talaromyces marneffei (formerly known as Penicillium marneffei). The majority of these organisms are primary pathogens, with the ability to cause disease in healthy humans who encounter them in endemic areas.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Abuodeh RO, Orbach MJ, Mandel MA, Das A, Galgiani JN. 2000. Genetic transformation of Coccidioides immitis facilitated by Agrobacterium tumefaciens. J Infect Dis 181: 2106–2110. - PubMed
-
- Albuquerque PC, Nakayasu ES, Rodrigues ML, Frases S, Casadevall A, Zancope-Oliveira RM, Almeida IC, Nosanchuk JD. 2008. Vesicular transport in Histoplasma capsulatum: An effective mechanism for trans-cell wall transfer of proteins and lipids in ascomycetes. Cell Microbiol 10: 1695–1710. - PMC - PubMed
-
- Almeida AJ, Carmona JA, Cunha C, Carvalho A, Rappleye CA, Goldman WE, Hooykaas PJ, Leao C, Ludovico P, Rodrigues F. 2007. Towards a molecular genetic system for the pathogenic fungus Paracoccidioides brasiliensis. Fungal Genet Biol 44: 1387–1398. - PubMed
-
- Almeida AJ, Cunha C, Carmona JA, Sampaio-Marques B, Carvalho A, Malavazi I, Steensma HY, Johnson DI, Leao C, Logarinho E, et al. 2009. Cdc42p controls yeast-cell shape and virulence of Paracoccidioides brasiliensis. Fungal Genet Biol 46: 919–926. - PubMed
-
- Andrade RV, Paes HC, Nicola AM, de Carvalho MJ, Fachin AL, Cardoso RS, Silva SS, Fernandes L, Silva SP, Donadi EA, et al. 2006. Cell organisation, sulphur metabolism and ion transport-related genes are differentially expressed in Paracoccidioides brasiliensis mycelium and yeast cells. BMC Genomics 7: 208. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical