

Human monocyte recognition of adenosine-based cyclic dinucleotides unveils the A2a Gαs protein-coupled receptor tonic inhibition of mitochondrially induced cell death
- PMID: 25384972
- PMCID: PMC4272430
- DOI: 10.1128/MCB.01204-14
Human monocyte recognition of adenosine-based cyclic dinucleotides unveils the A2a Gαs protein-coupled receptor tonic inhibition of mitochondrially induced cell death
Abstract
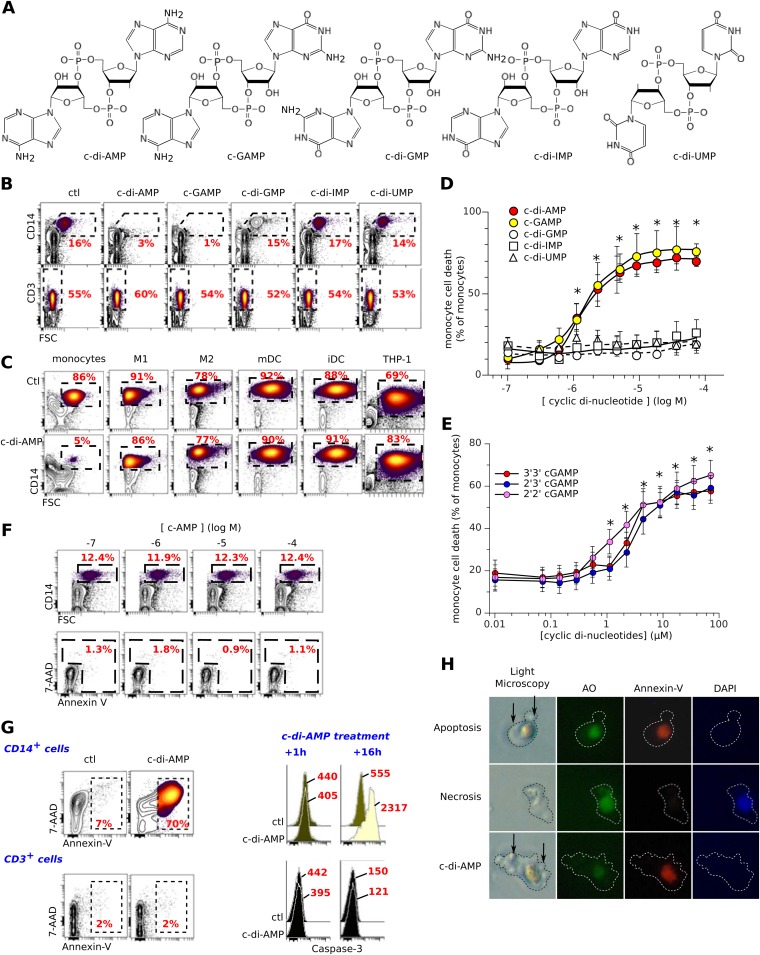
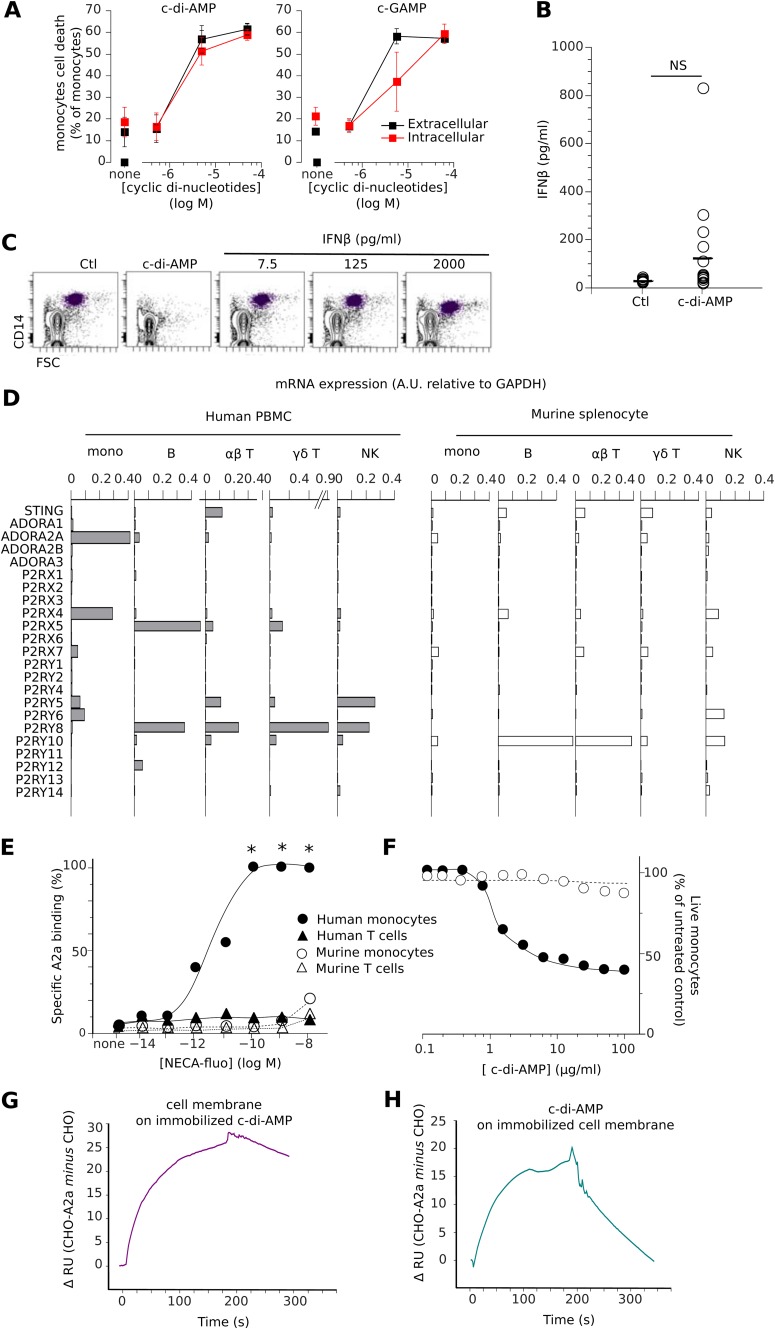
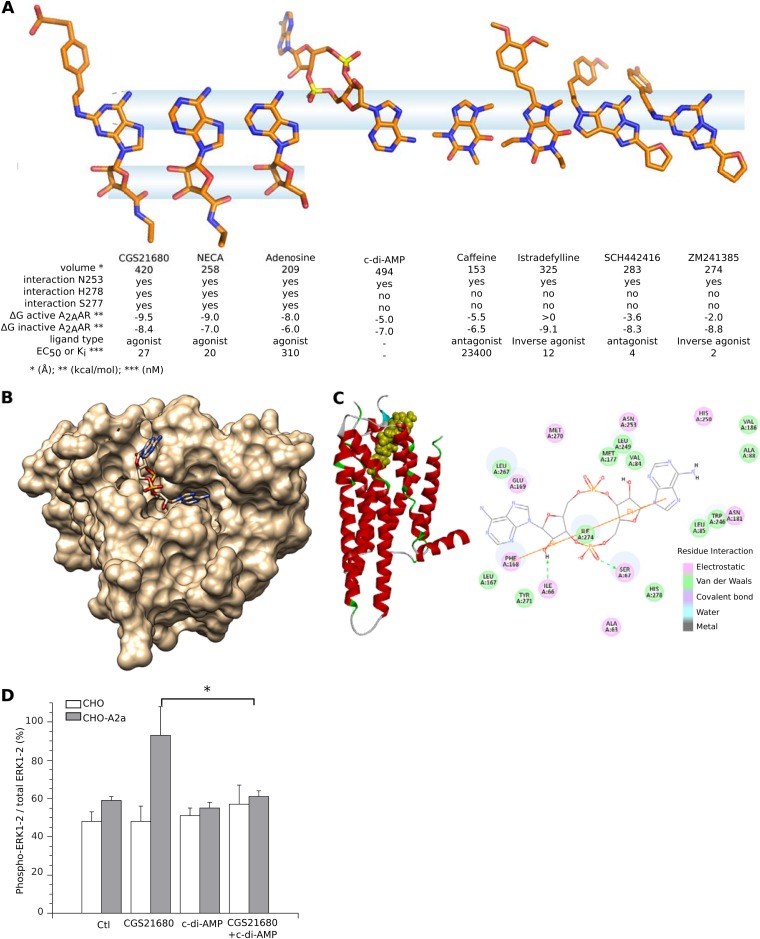
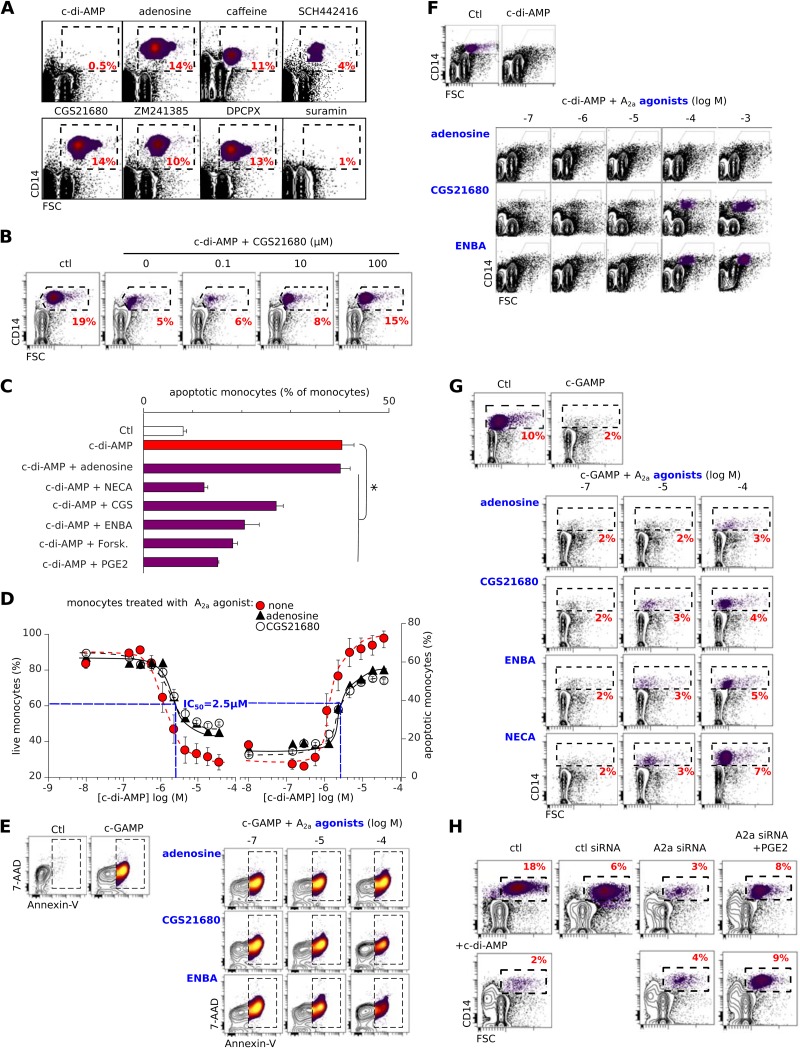
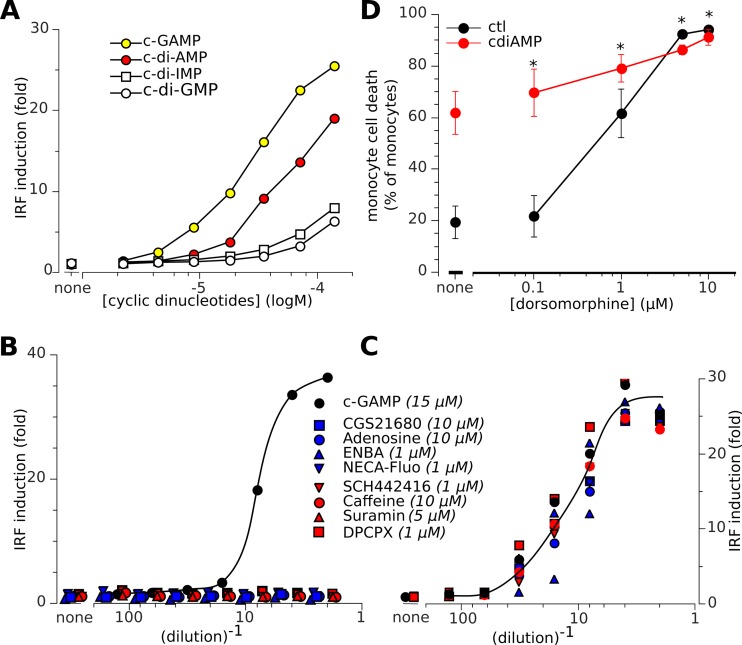
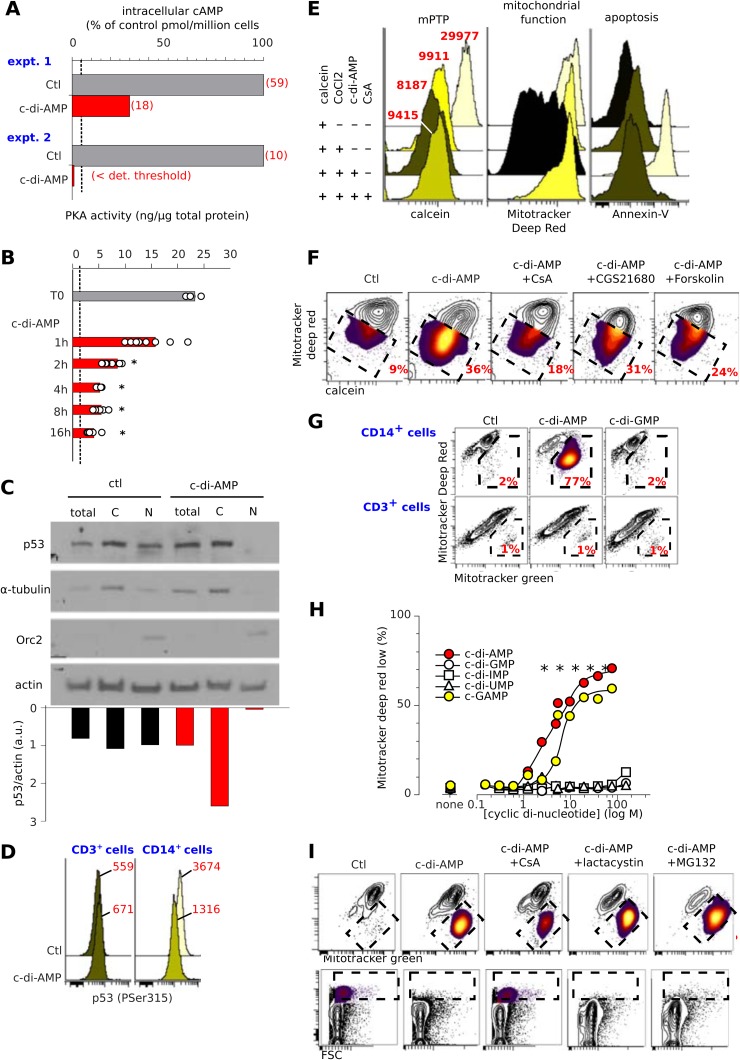
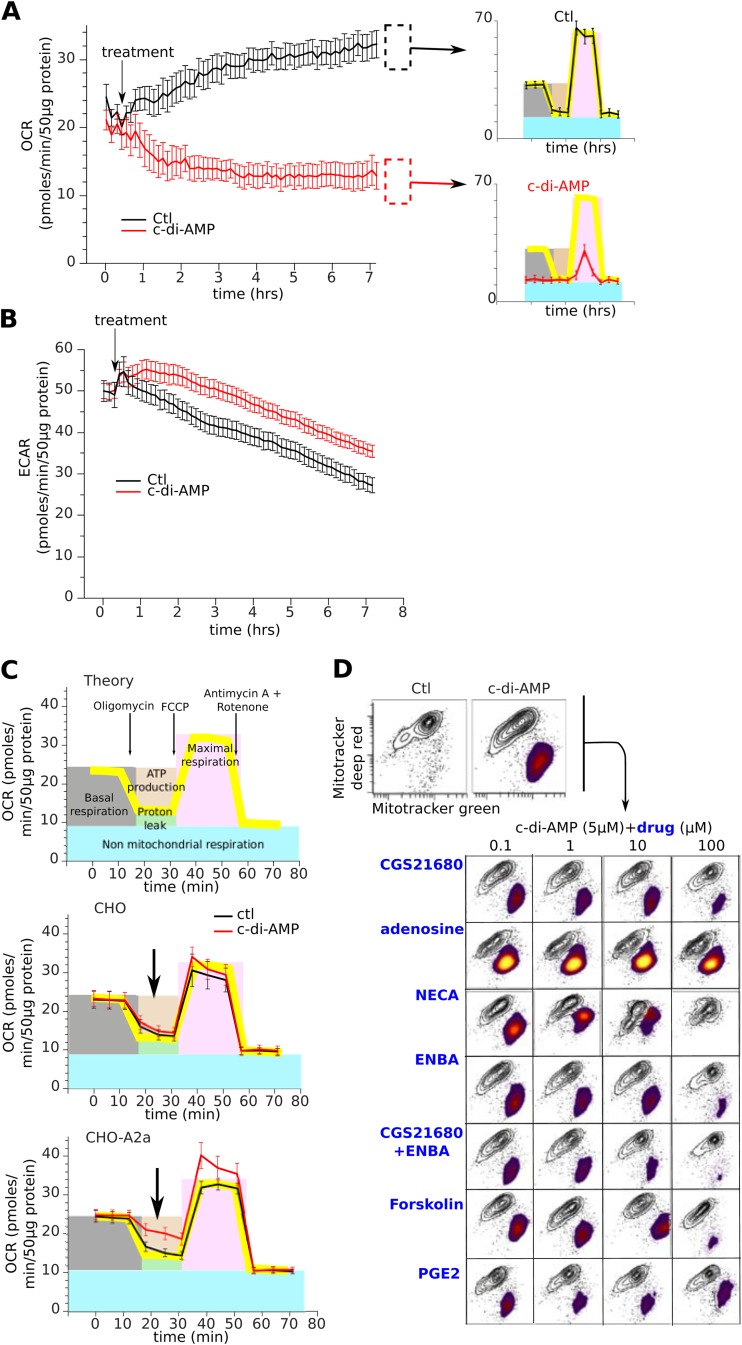
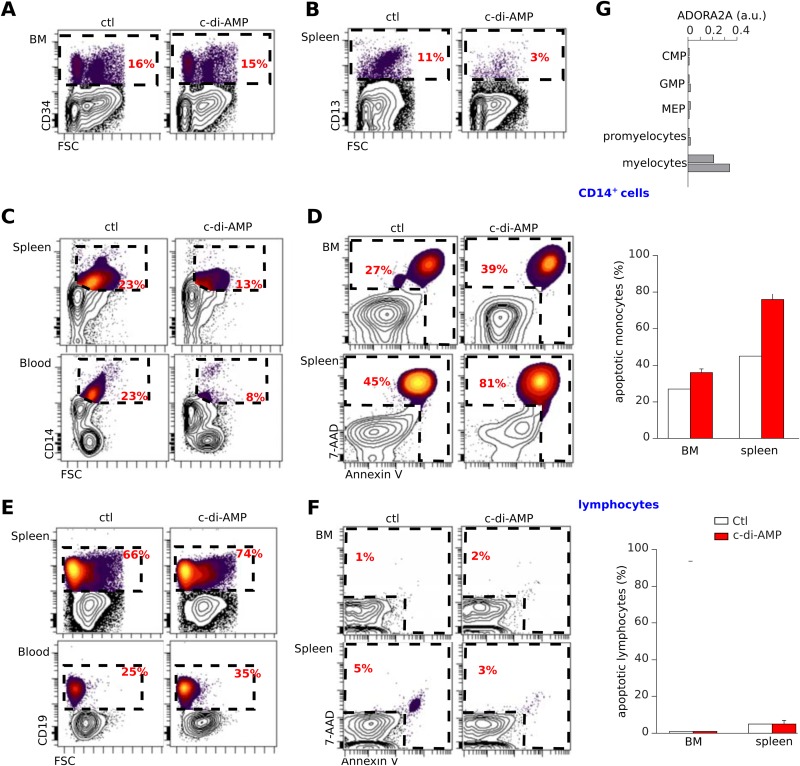
Cyclic dinucleotides are important messengers for bacteria and protozoa and are well-characterized immunity alarmins for infected mammalian cells through intracellular binding to STING receptors. We sought to investigate their unknown extracellular effects by adding cyclic dinucleotides to the culture medium of freshly isolated human blood cells in vitro. Here we report that adenosine-containing cyclic dinucleotides induce the selective apoptosis of monocytes through a novel apoptotic pathway. We demonstrate that these compounds are inverse agonist ligands of A2a, a Gαs-coupled adenosine receptor selectively expressed by monocytes. Inhibition of monocyte A2a by these ligands induces apoptosis through a mechanism independent of that of the STING receptors. The blockade of basal (adenosine-free) signaling from A2a inhibits protein kinase A (PKA) activity, thereby recruiting cytosolic p53, which opens the mitochondrial permeability transition pore and impairs mitochondrial respiration, resulting in apoptosis. A2a antagonists and inverse agonist ligands induce apoptosis of human monocytes, while A2a agonists are antiapoptotic. In vivo, we used a mock developing human hematopoietic system through NSG mice transplanted with human CD34(+) cells. Treatment with cyclic di-AMP selectively depleted A2a-expressing monocytes and their precursors via apoptosis. Thus, monocyte recognition of cyclic dinucleotides unravels a novel proapoptotic pathway: the A2a Gαs protein-coupled receptor (GPCR)-driven tonic inhibitory signaling of mitochondrion-induced cell death.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous