

Alzheimer's therapeutics targeting amyloid beta 1-42 oligomers II: Sigma-2/PGRMC1 receptors mediate Abeta 42 oligomer binding and synaptotoxicity
- PMID: 25390692
- PMCID: PMC4229119
- DOI: 10.1371/journal.pone.0111899
Alzheimer's therapeutics targeting amyloid beta 1-42 oligomers II: Sigma-2/PGRMC1 receptors mediate Abeta 42 oligomer binding and synaptotoxicity
Abstract
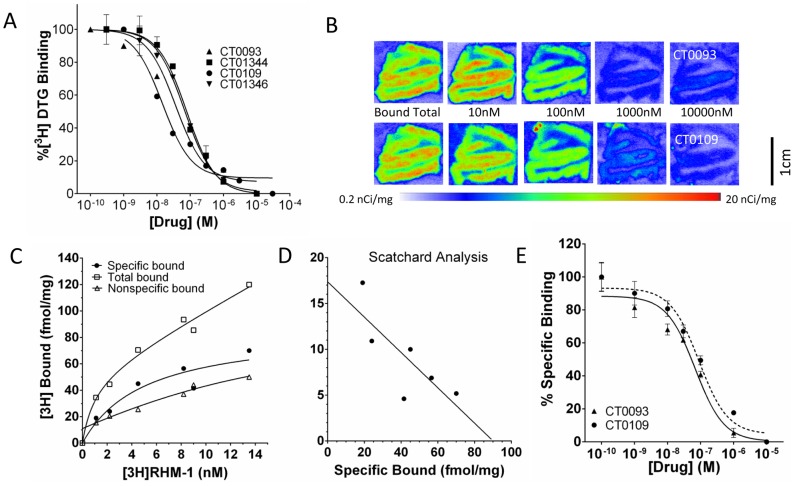
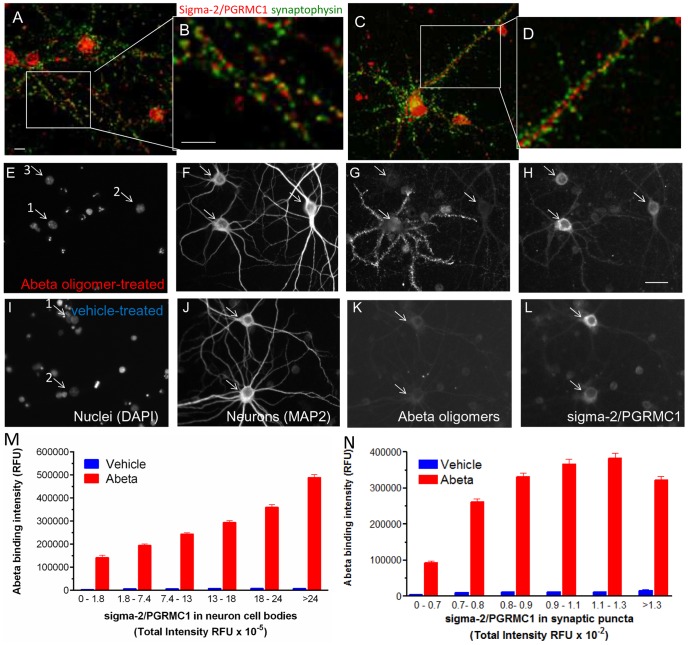
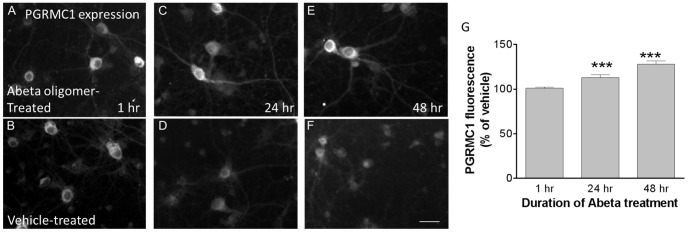
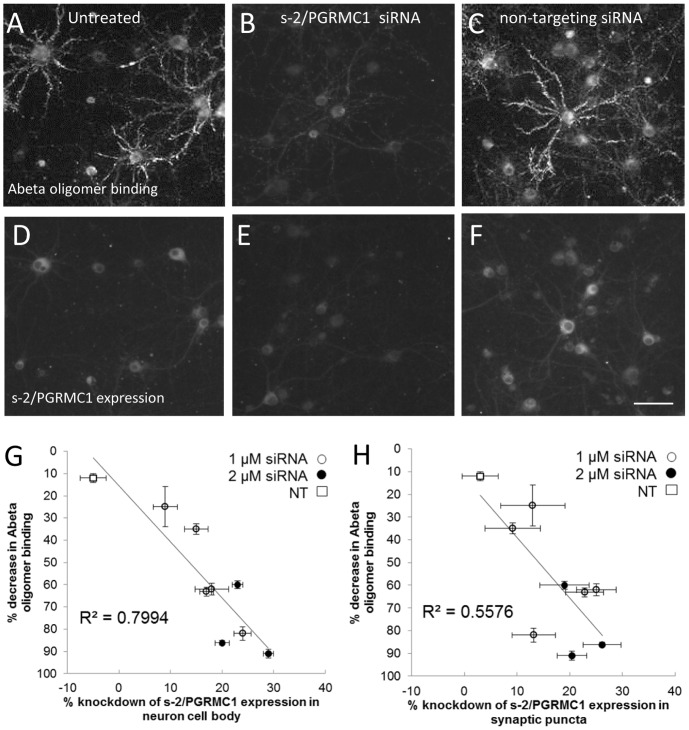
Amyloid beta (Abeta) 1-42 oligomers accumulate in brains of patients with Mild Cognitive Impairment (MCI) and disrupt synaptic plasticity processes that underlie memory formation. Synaptic binding of Abeta oligomers to several putative receptor proteins is reported to inhibit long-term potentiation, affect membrane trafficking and induce reversible spine loss in neurons, leading to impaired cognitive performance and ultimately to anterograde amnesia in the early stages of Alzheimer's disease (AD). We have identified a receptor not previously associated with AD that mediates the binding of Abeta oligomers to neurons, and describe novel therapeutic antagonists of this receptor capable of blocking Abeta toxic effects on synapses in vitro and cognitive deficits in vivo. Knockdown of sigma-2/PGRMC1 (progesterone receptor membrane component 1) protein expression in vitro using siRNA results in a highly correlated reduction in binding of exogenous Abeta oligomers to neurons of more than 90%. Expression of sigma-2/PGRMC1 is upregulated in vitro by treatment with Abeta oligomers, and is dysregulated in Alzheimer's disease patients' brain compared to age-matched, normal individuals. Specific, high affinity small molecule receptor antagonists and antibodies raised against specific regions on this receptor can displace synthetic Abeta oligomer binding to synaptic puncta in vitro and displace endogenous human AD patient oligomers from brain tissue sections in a dose-dependent manner. These receptor antagonists prevent and reverse the effects of Abeta oligomers on membrane trafficking and synapse loss in vitro and cognitive deficits in AD mouse models. These findings suggest sigma-2/PGRMC1 receptors mediate saturable oligomer binding to synaptic puncta on neurons and that brain penetrant, small molecules can displace endogenous and synthetic oligomers and improve cognitive deficits in AD models. We propose that sigma-2/PGRMC1 is a key mediator of the pathological effects of Abeta oligomers in AD and is a tractable target for small molecule disease-modifying therapeutics.
Conflict of interest statement
Figures
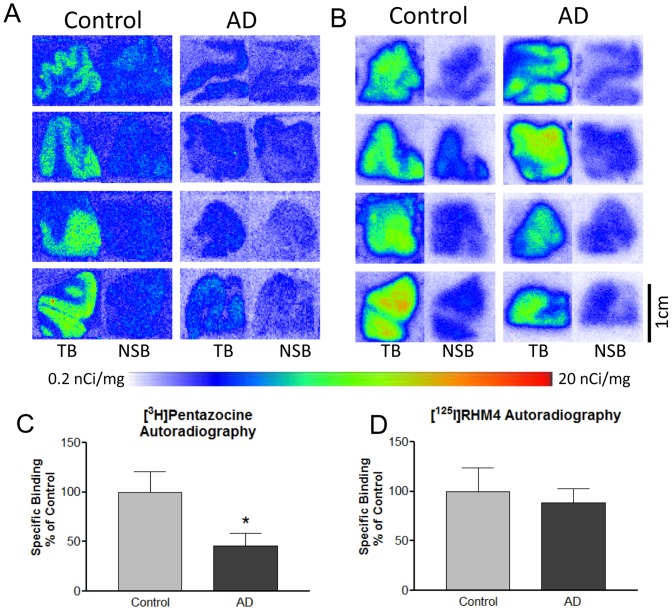
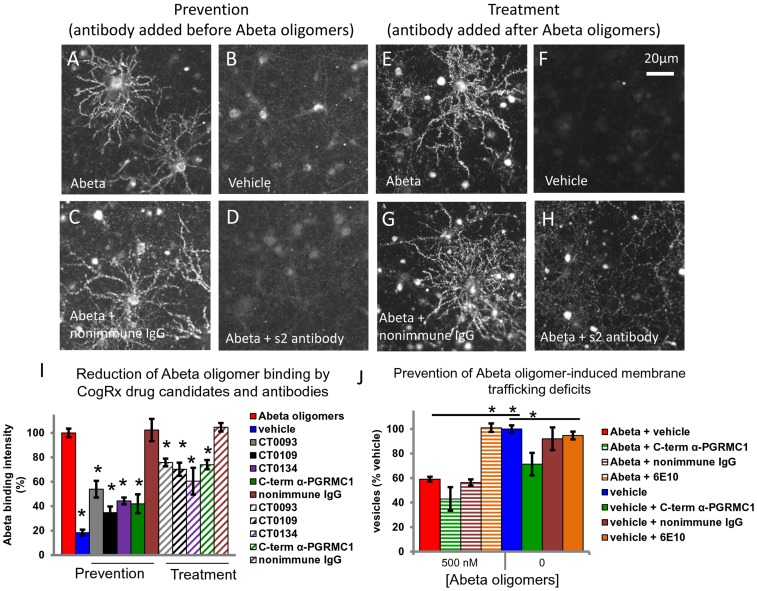
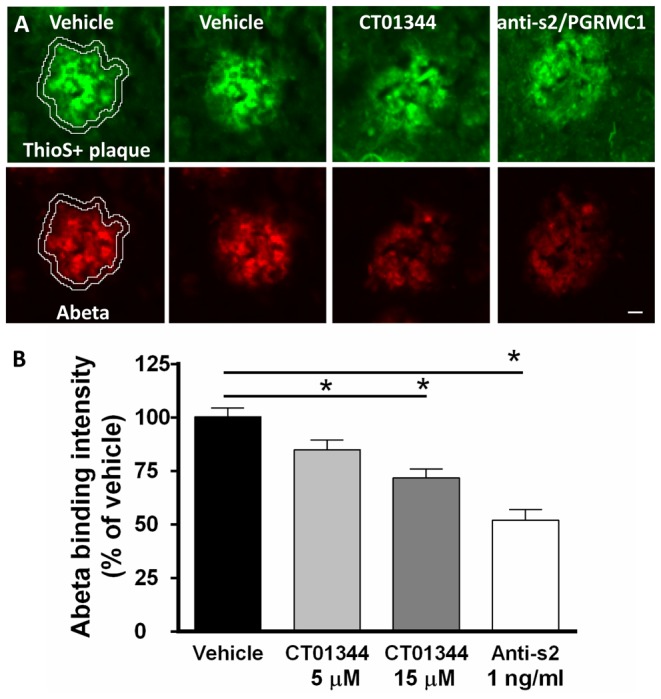
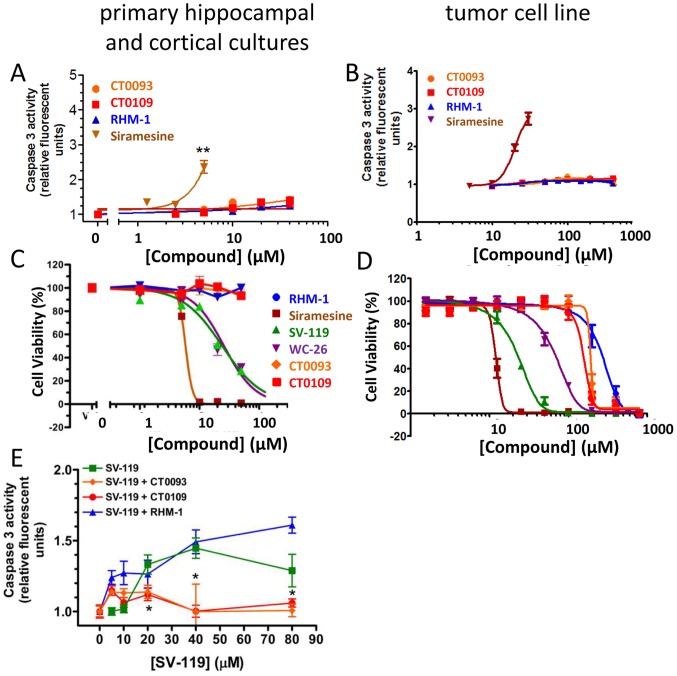
References
-
- Klein WL, Krafft G, Finch CE (2001) Targeting small Abeta oligomers: the solution to an Alzheimer's disease conundrum. Trends Neurosci 24: 219–224. - PubMed
-
- Bao F, Wicklund L, Lacor PN, Klein WL, Nordberg A, et al. (2012) Different beta-amyloid oligomer assemblies in Alzheimer brains correlate with age of disease onset and impaired cholinergic activity. Neurobiol Aging 33: 813 –813. - PubMed
-
- Haes AJ, Chang L, Klein WL, Van Duyne RP (2005) Detection of a biomarker for Alzheimer's disease from synthetic and clinical samples using a nanoscale optical biosensor. J Am Chem Soc 127: 2264–2271. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
