

Selective inhibition of mutant isocitrate dehydrogenase 1 (IDH1) via disruption of a metal binding network by an allosteric small molecule
- PMID: 25391653
- PMCID: PMC4294499
- DOI: 10.1074/jbc.M114.608497
Selective inhibition of mutant isocitrate dehydrogenase 1 (IDH1) via disruption of a metal binding network by an allosteric small molecule
Abstract
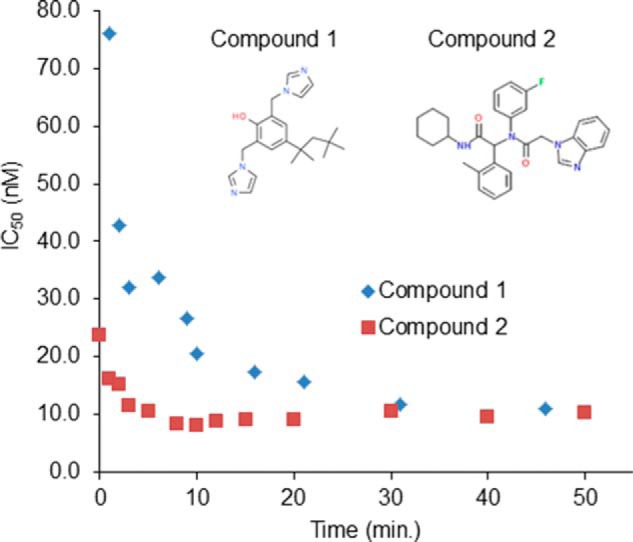
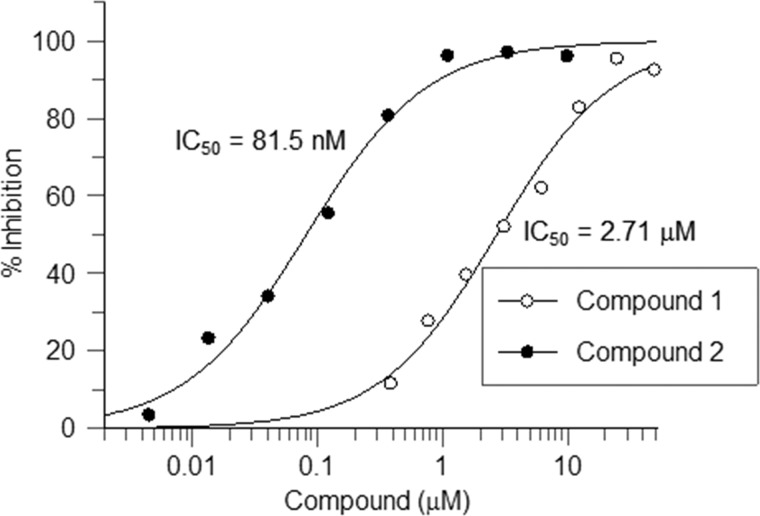
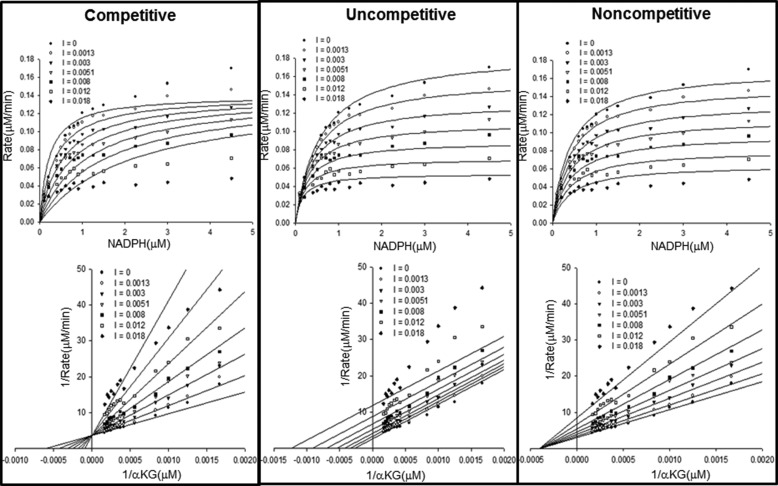
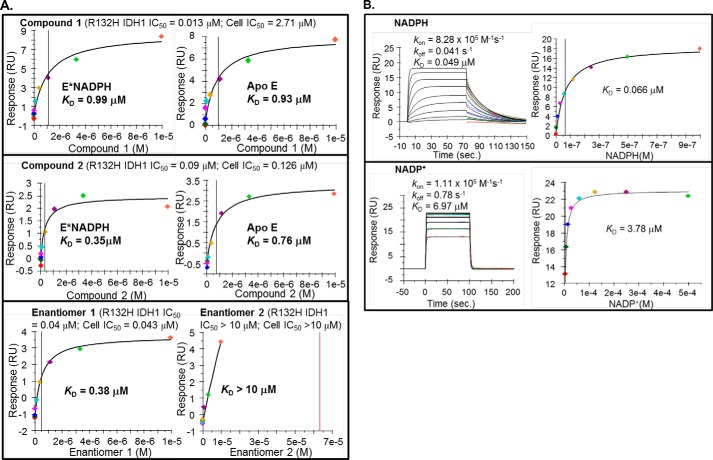
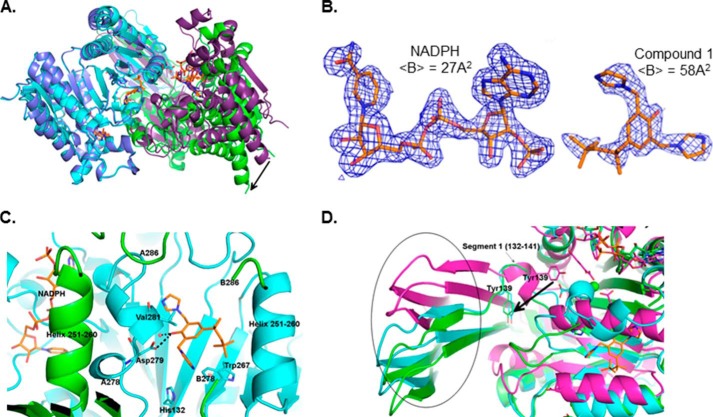
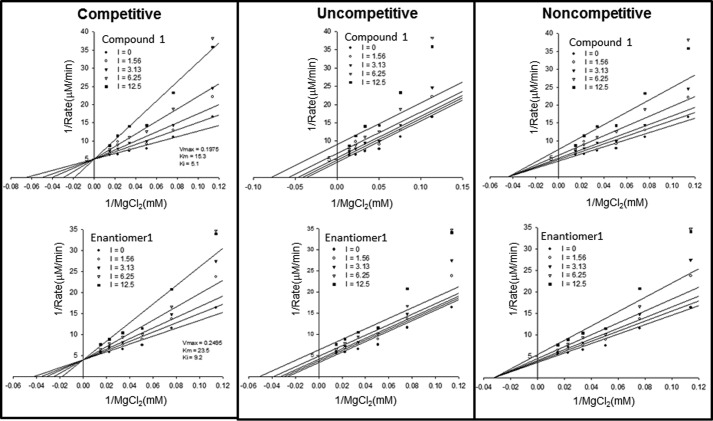
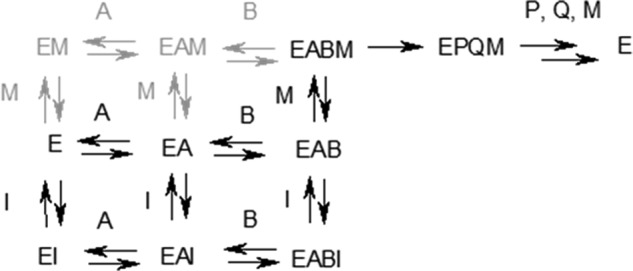
Cancer-associated point mutations in isocitrate dehydrogenase 1 and 2 (IDH1 and IDH2) confer a neomorphic enzymatic activity: the reduction of α-ketoglutarate to d-2-hydroxyglutaric acid, which is proposed to act as an oncogenic metabolite by inducing hypermethylation of histones and DNA. Although selective inhibitors of mutant IDH1 and IDH2 have been identified and are currently under investigation as potential cancer therapeutics, the mechanistic basis for their selectivity is not yet well understood. A high throughput screen for selective inhibitors of IDH1 bearing the oncogenic mutation R132H identified compound 1, a bis-imidazole phenol that inhibits d-2-hydroxyglutaric acid production in cells. We investigated the mode of inhibition of compound 1 and a previously published IDH1 mutant inhibitor with a different chemical scaffold. Steady-state kinetics and biophysical studies show that both of these compounds selectively inhibit mutant IDH1 by binding to an allosteric site and that inhibition is competitive with respect to Mg(2+). A crystal structure of compound 1 complexed with R132H IDH1 indicates that the inhibitor binds at the dimer interface and makes direct contact with a residue involved in binding of the catalytically essential divalent cation. These results show that targeting a divalent cation binding residue can enable selective inhibition of mutant IDH1 and suggest that differences in magnesium binding between wild-type and mutant enzymes may contribute to the inhibitors' selectivity for the mutant enzyme.
Keywords: Anticancer Drug; Cancer Metabolism; Drug Action; Drug Discovery; Enzyme Inhibitor; IDH1 Inhibitor; Mechanism of Inhibition; Metalloenzyme.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Tennant D. A., Durán R. V., Gottlieb E. (2010) Targeting metabolic transformation for cancer therapy. Nat. Rev. Cancer 10, 267–277 - PubMed
-
- Levine A. J., Puzio-Kuter A. M. (2010) The control of the metabolic switch in cancers by oncogenes and tumor suppressor genes. Science 330, 1340–1344 - PubMed
-
- Dang L., Jin S., Su S. M. (2010) IDH mutations in glioma and acute myeloid leukemia. Trends Mol. Med. 16, 387–397 - PubMed
-
- Geisbrecht B. V., Gould S. J. (1999) The human PICD gene encodes a cytoplasmic and peroxisomal NADP+-dependent isocitrate dehydrogenase. J. Biol. Chem. 274, 30527–30533 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
