

Epo-induced erythroid maturation is dependent on Plcγ1 signaling
- PMID: 25394487
- PMCID: PMC4423181
- DOI: 10.1038/cdd.2014.186
Epo-induced erythroid maturation is dependent on Plcγ1 signaling
Abstract
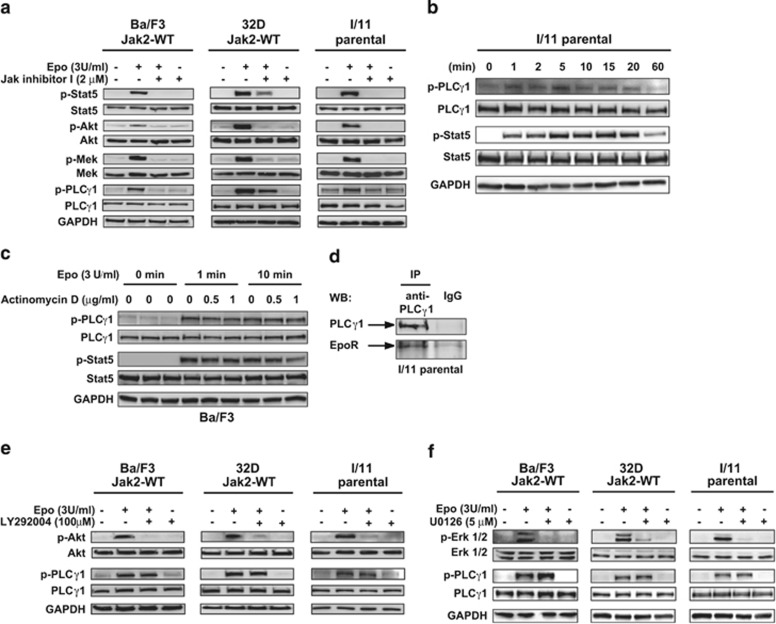
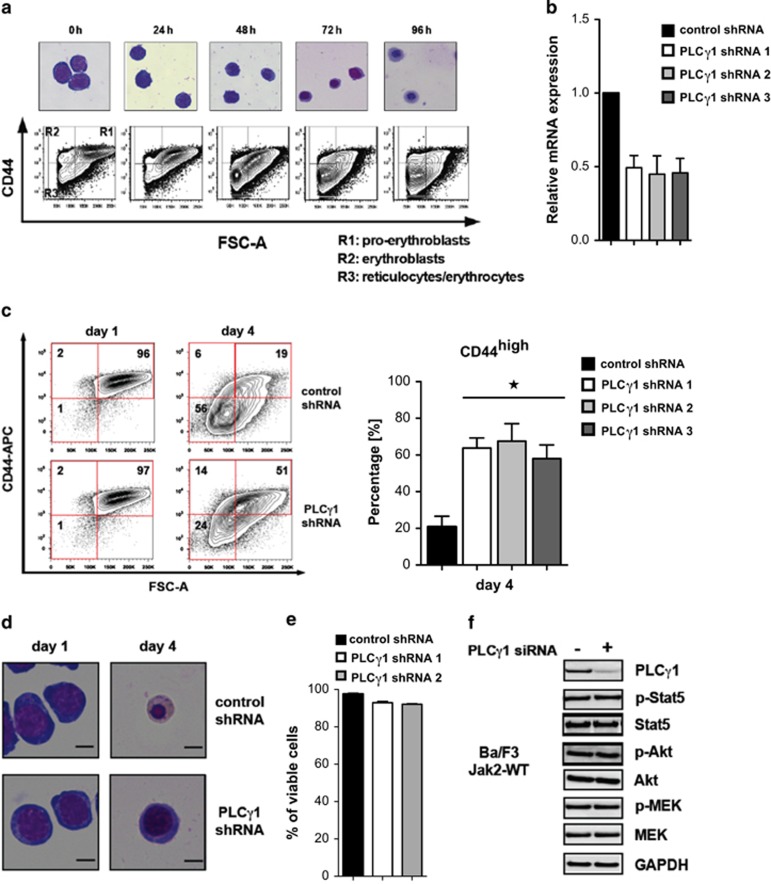
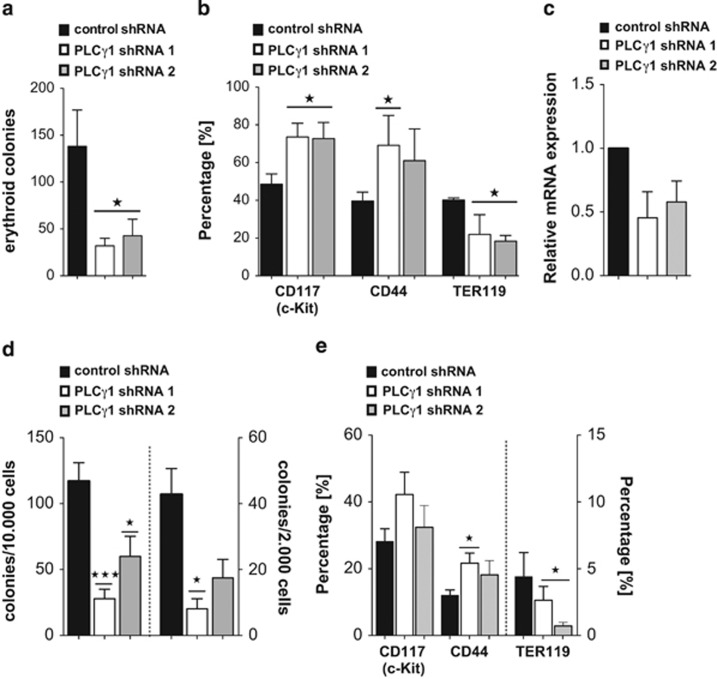
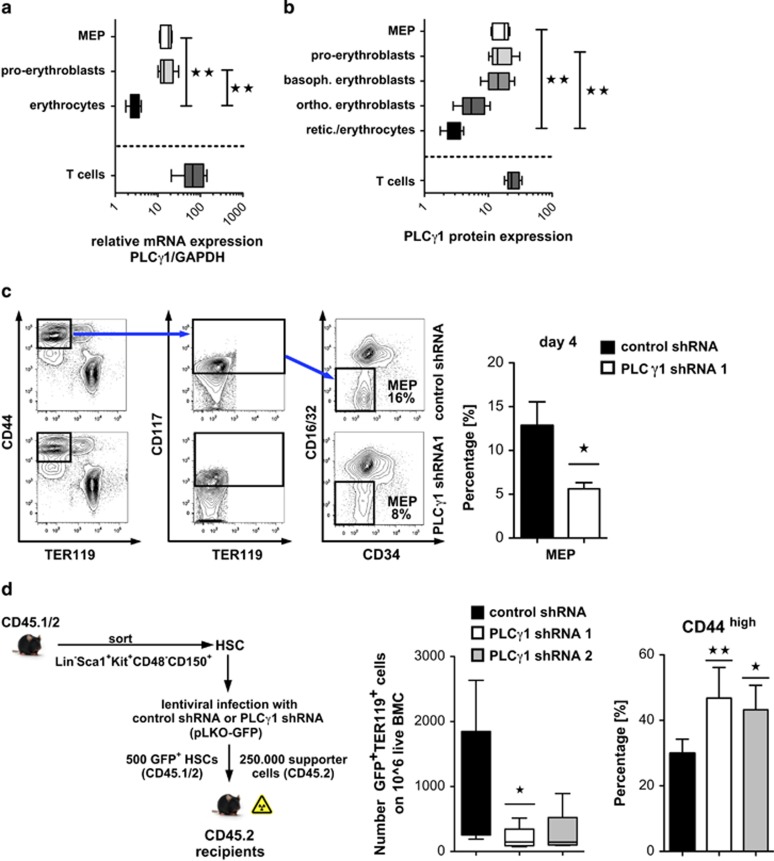
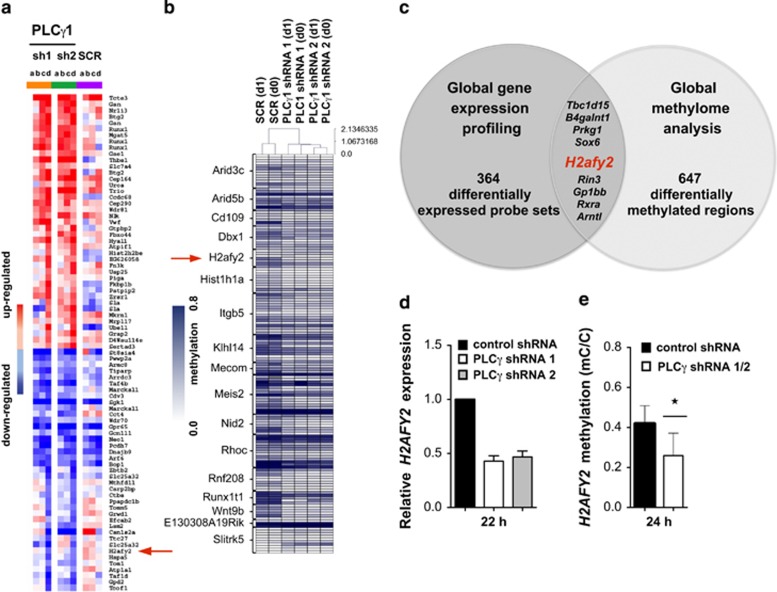
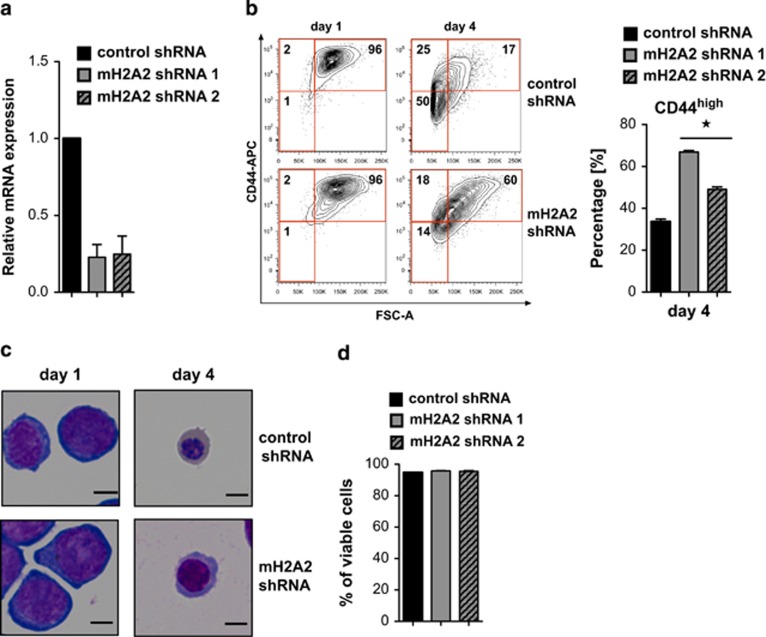
Erythropoiesis is a tightly regulated process. Development of red blood cells occurs through differentiation of hematopoietic stem cells (HSCs) into more committed progenitors and finally into erythrocytes. Binding of erythropoietin (Epo) to its receptor (EpoR) is required for erythropoiesis as it promotes survival and late maturation of erythroid progenitors. In vivo and in vitro studies have highlighted the requirement of EpoR signaling through Janus kinase 2 (Jak2) tyrosine kinase and Stat5a/b as a central pathway. Here, we demonstrate that phospholipase C gamma 1 (Plcγ1) is activated downstream of EpoR-Jak2 independently of Stat5. Plcγ1-deficient pro-erythroblasts and erythroid progenitors exhibited strong impairment in differentiation and colony-forming potential. In vivo, suppression of Plcγ1 in immunophenotypically defined HSCs (Lin(-)Sca1(+)KIT(+)CD48(-)CD150(+)) severely reduced erythroid development. To identify Plcγ1 effector molecules involved in regulation of erythroid differentiation, we assessed changes occurring at the global transcriptional and DNA methylation level after inactivation of Plcγ1. The top common downstream effector was H2afy2, which encodes for the histone variant macroH2A2 (mH2A2). Inactivation of mH2A2 expression recapitulated the effects of Plcγ1 depletion on erythroid maturation. Taken together, our findings identify Plcγ1 and its downstream target mH2A2, as a 'non-canonical' Epo signaling pathway essential for erythroid differentiation.
Figures
References
-
- Richmond TD, Chohan M, Barber DL. Turning cells red: signal transduction mediated by erythropoietin. Trends Cell Biol. 2005;15:146–155. - PubMed
-
- Paulson RF, Bernstein A. Receptor tyrosine kinases and the regulation of hematopoiesis. Semin Immunol. 1995;7:267–277. - PubMed
-
- Cantor AB, Orkin SH. Hematopoietic development: a balancing act. Curr Opin Genet Dev. 2001;11:513–519. - PubMed
-
- Wu H, Liu X, Jaenisch R, Lodish HF. Generation of committed erythroid BFU-E and CFU-E progenitors does not require erythropoietin or the erythropoietin receptor. Cell. 1995;83:59–67. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
