

CSN6 drives carcinogenesis by positively regulating Myc stability
- PMID: 25395170
- PMCID: PMC4234183
- DOI: 10.1038/ncomms6384
CSN6 drives carcinogenesis by positively regulating Myc stability
Abstract
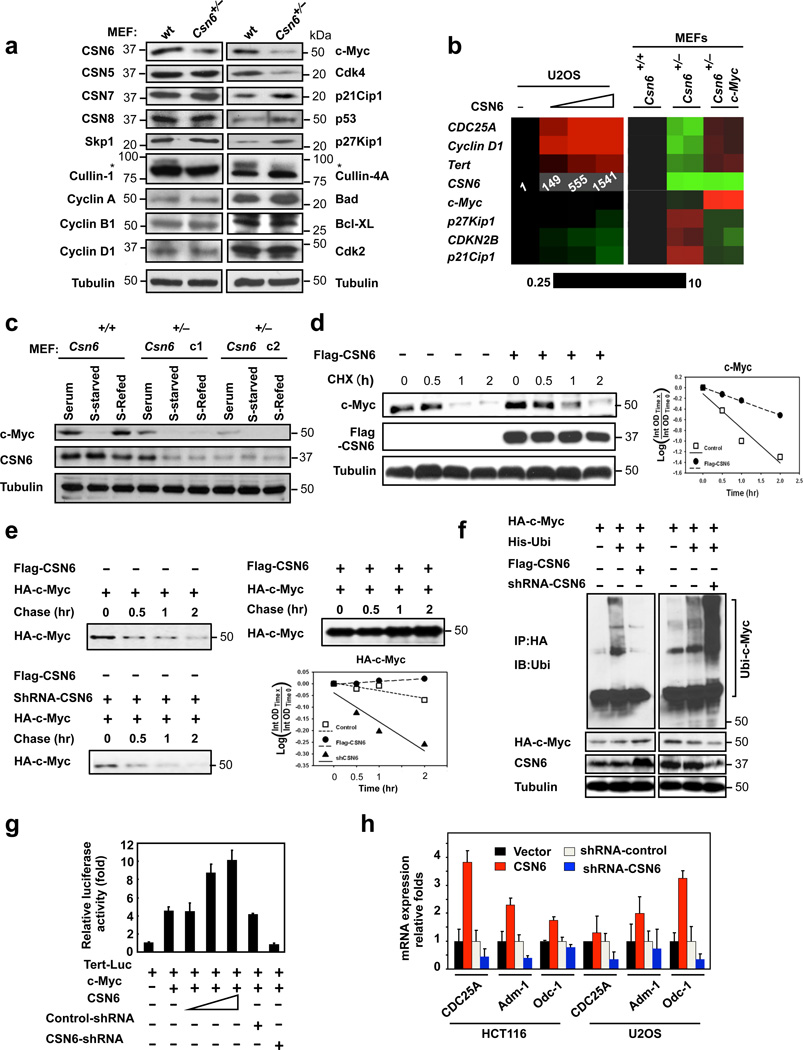
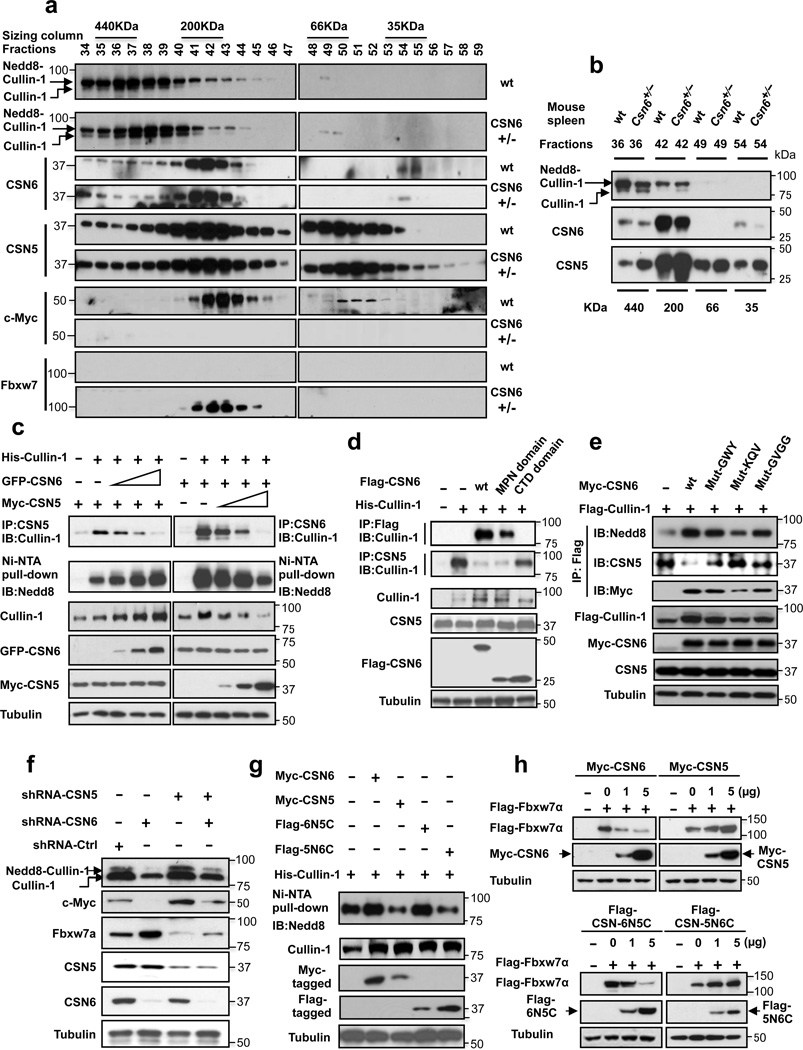
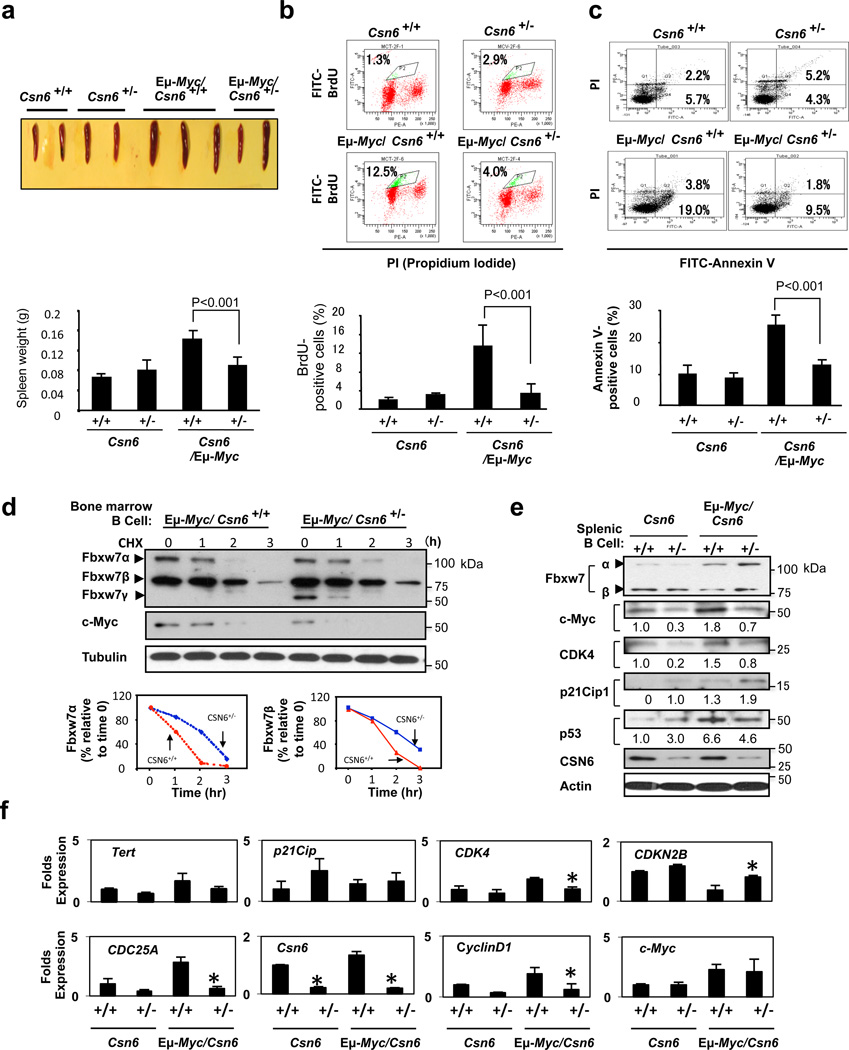
Cullin-RING ubiquitin ligases (CRLs) are critical in ubiquitinating Myc, while COP9 signalosome (CSN) controls neddylation of Cullin in CRL. The mechanistic link between Cullin neddylation and Myc ubiquitination/degradation is unclear. Here we show that Myc is a target of the CSN subunit 6 (CSN6)-Cullin signalling axis and that CSN6 is a positive regulator of Myc. CSN6 enhanced neddylation of Cullin-1 and facilitated autoubiquitination/degradation of Fbxw7, a component of CRL involved in Myc ubiquitination, thereby stabilizing Myc. Csn6 haplo-insufficiency decreased Cullin-1 neddylation but increased Fbxw7 stability to compromise Myc stability and activity in an Eμ-Myc mouse model, resulting in decelerated lymphomagenesis. We found that CSN6 overexpression, which leads to aberrant expression of Myc target genes, is frequent in human cancers. Together, these results define a mechanism for the regulation of Myc stability through the CSN-Cullin-Fbxw7 axis and provide insights into the correlation of CSN6 overexpression with Myc stabilization/activation during tumorigenesis.
Figures




References
-
- Wei N, Deng XW. The COP9 signalosome. Annu Rev Cell Dev Biol. 2003;19:261–286. - PubMed
-
- Serino G, Deng XW. The COP9 signalosome: regulating plant development through the control of proteolysis. Annu Rev Plant Biol. 2003;54:165–182. - PubMed
-
- Chamovitz DA, et al. The COP9 complex, a novel multisubunit nuclear regulator involved in light control of a plant developmental switch. Cell. 1996;86:115–121. - PubMed
-
- Wei N, Deng XW. Making sense of the COP9 signalosome. A regulatory protein complex conserved from Arabidopsis to human. Trends Genet. 1999;15:98–103. - PubMed
-
- Lyapina S, et al. Promotion of NEDD-CUL1 conjugate cleavage by COP9 signalosome. Science. 2001;292:1382–1385. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
