

Chromosome congression is promoted by CENP-Q- and CENP-E-dependent pathways
- PMID: 25395579
- PMCID: PMC4282051
- DOI: 10.1242/jcs.163659
Chromosome congression is promoted by CENP-Q- and CENP-E-dependent pathways
Abstract
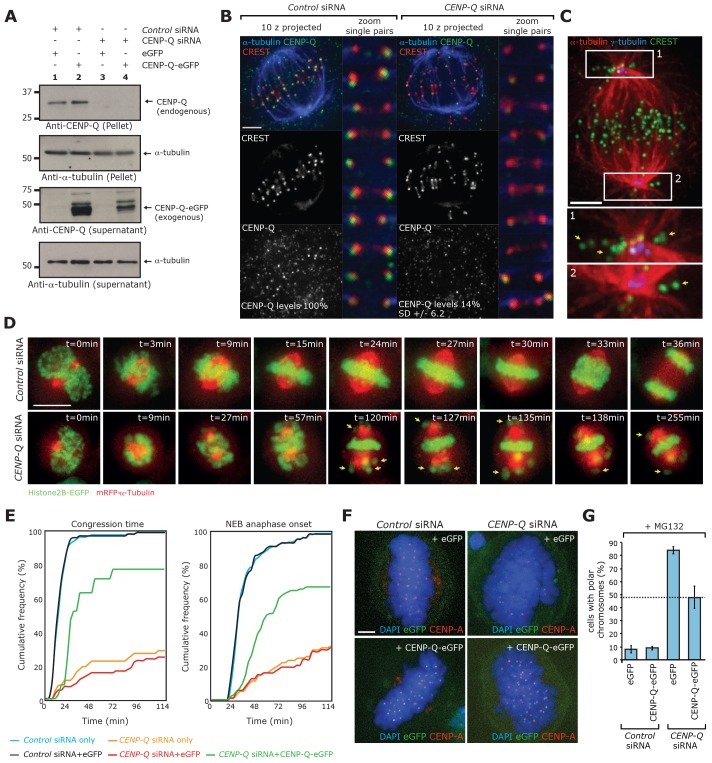
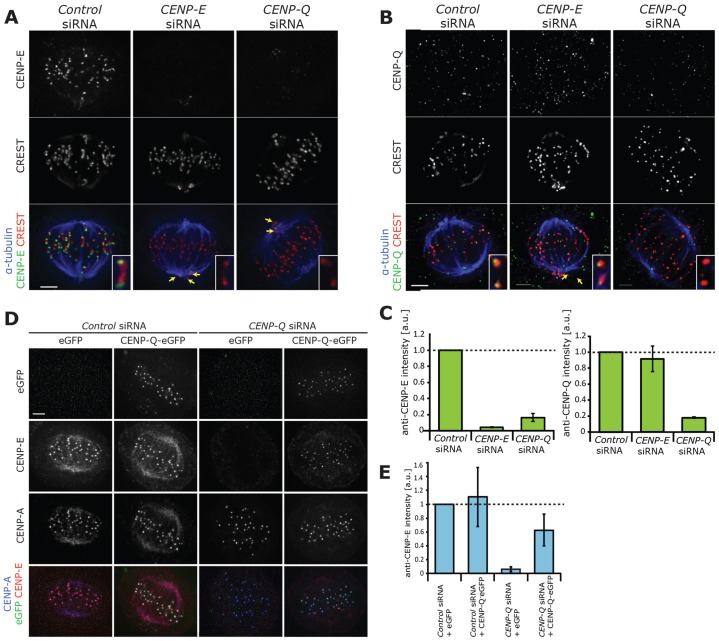
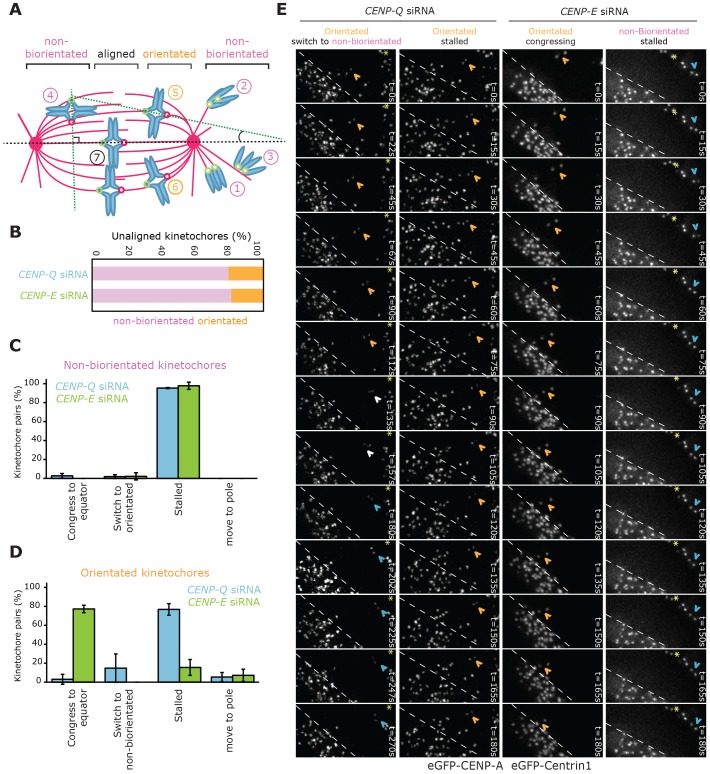
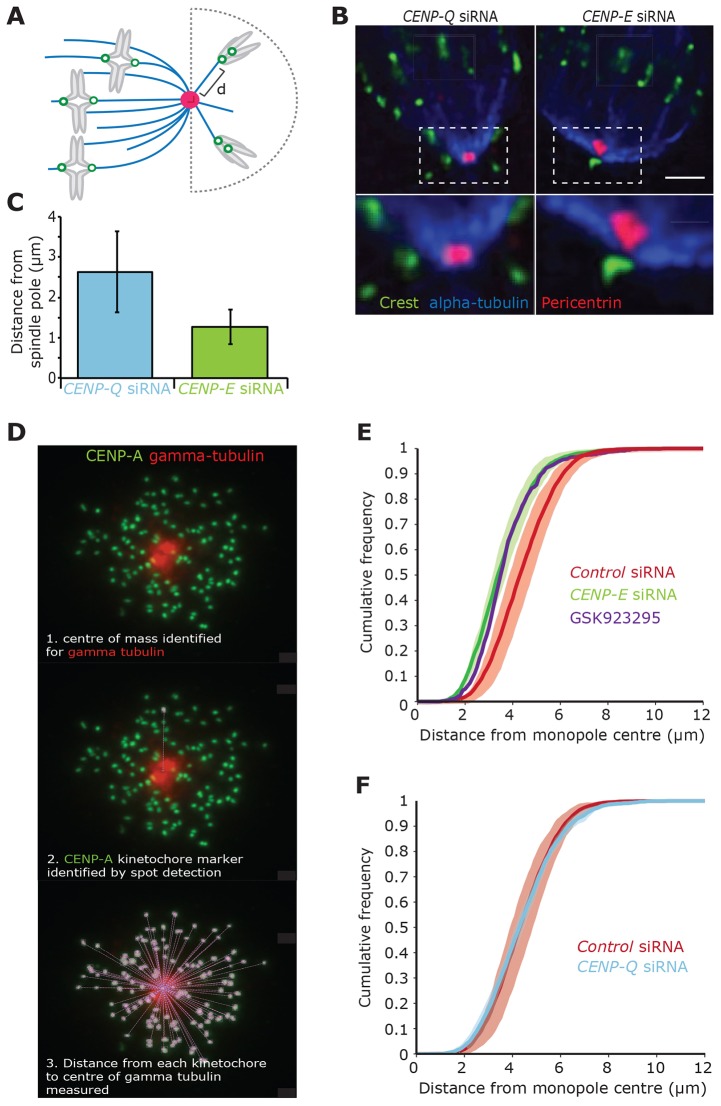
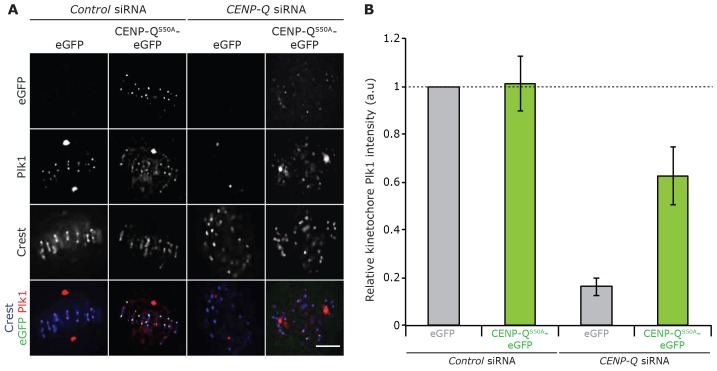
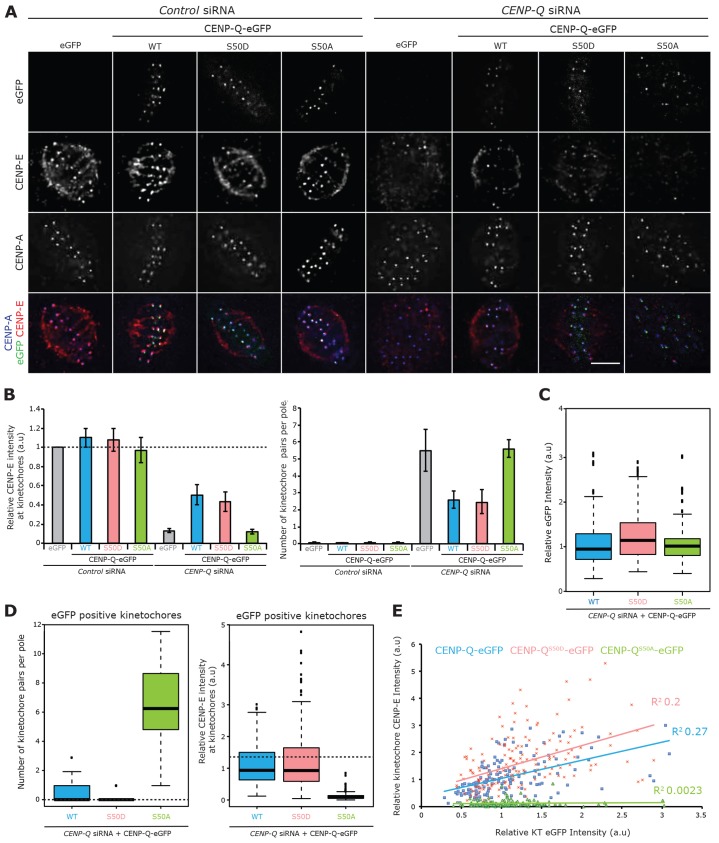
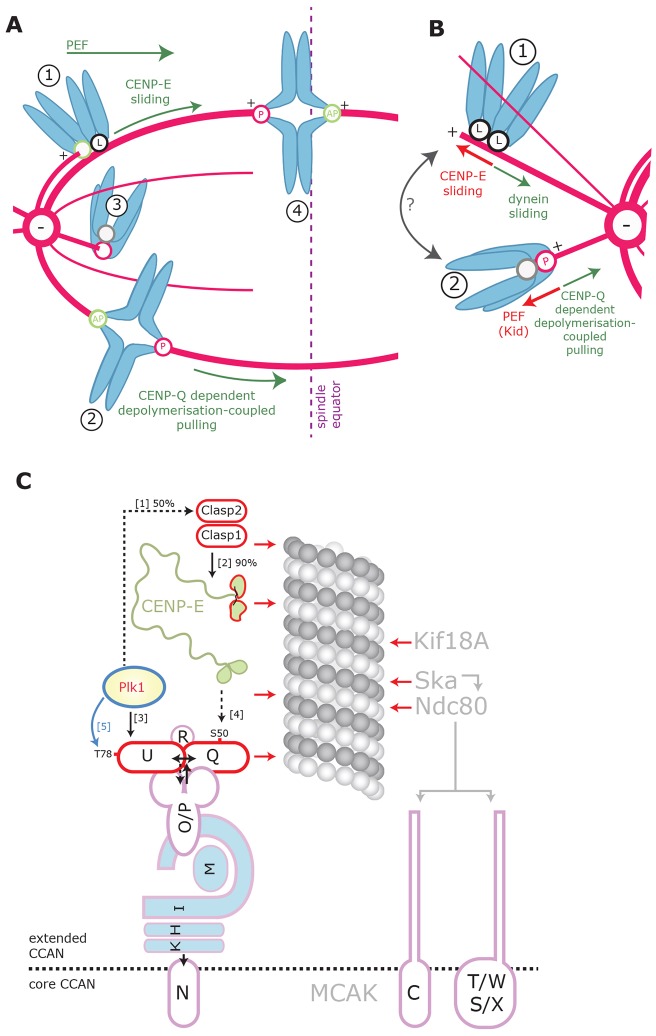
A key step of mitosis is the congression of chromosomes to the spindle equator. Congression is driven by at least two distinct mechanisms: (1) kinetochores slide along the microtubule lattice using the plus-end directed CENP-E motor, and (2) kinetochores biorientating near the pole move to the equator through microtubule depolymerisation-coupled pulling. Here, we show that CENP-Q - a subunit of the CENP-O complex (comprising CENP-O, CENP-P, CENP-Q and CENP-U) that targets polo-like kinase (Plk1) to kinetochores - is also required for the recruitment of CENP-E to kinetochores. We further reveal a CENP-E recruitment-independent role for CENP-Q in depolymerisation-coupled pulling. Both of these functions are abolished by a single point mutation in CENP-Q (S50A) - a residue that is phosphorylated in vivo. Importantly, the S50A mutant does not affect the loading of Plk1 onto kinetochores and leaves the CENP-O complex intact. Thus, the functions of CENP-Q in CENP-E loading and depolymerisation-coupled pulling are independent from its role in Plk1 recruitment and CENP-O complex stabilisation. Taken together, our data provide evidence that phosphoregulation of CENP-Q plays a central function in coordinating chromosome congression mechanisms.
Keywords: CENP-E; CENP-Q; Congression; Kinetochore; Mitosis.
© 2015. Published by The Company of Biologists Ltd.
Figures
References
-
- Ahonen L. J., Kallio M. J., Daum J. R., Bolton M., Manke I. A., Yaffe M. B., Stukenberg P. T., Gorbsky G. J. (2005). Polo-like kinase 1 creates the tension-sensing 3F3/2 phosphoepitope and modulates the association of spindle-checkpoint proteins at kinetochores. Curr. Biol. 15, 1078–1089 10.1016/j.cub.2005.05.026 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
