

INPP5E interacts with AURKA, linking phosphoinositide signaling to primary cilium stability
- PMID: 25395580
- PMCID: PMC4294778
- DOI: 10.1242/jcs.161323
INPP5E interacts with AURKA, linking phosphoinositide signaling to primary cilium stability
Abstract
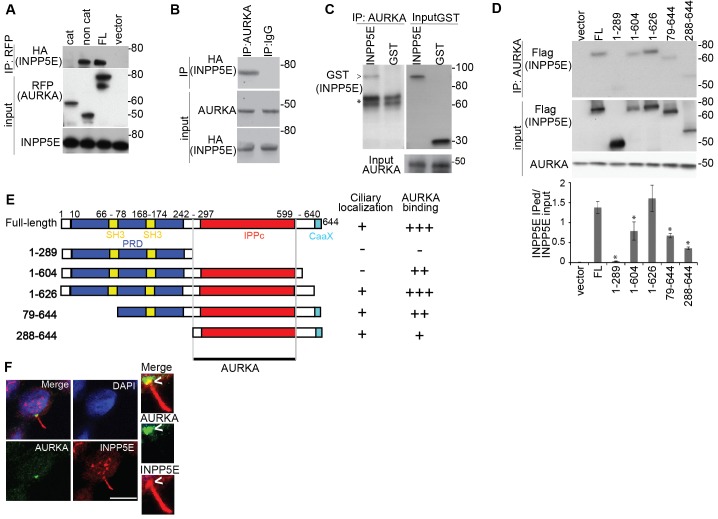
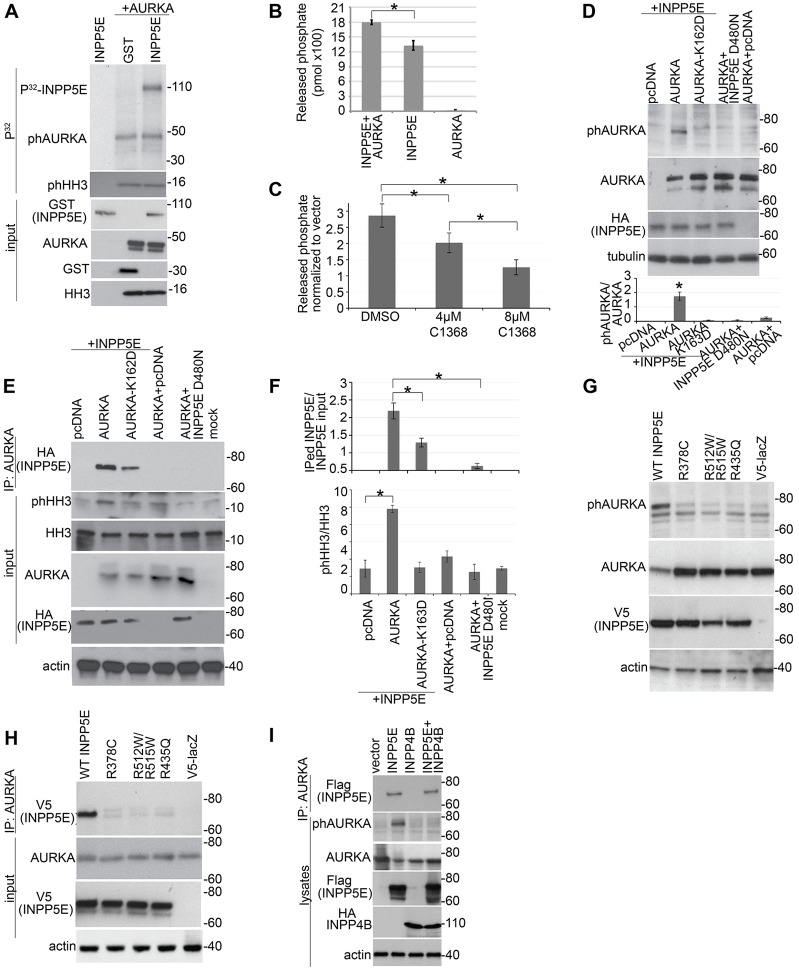
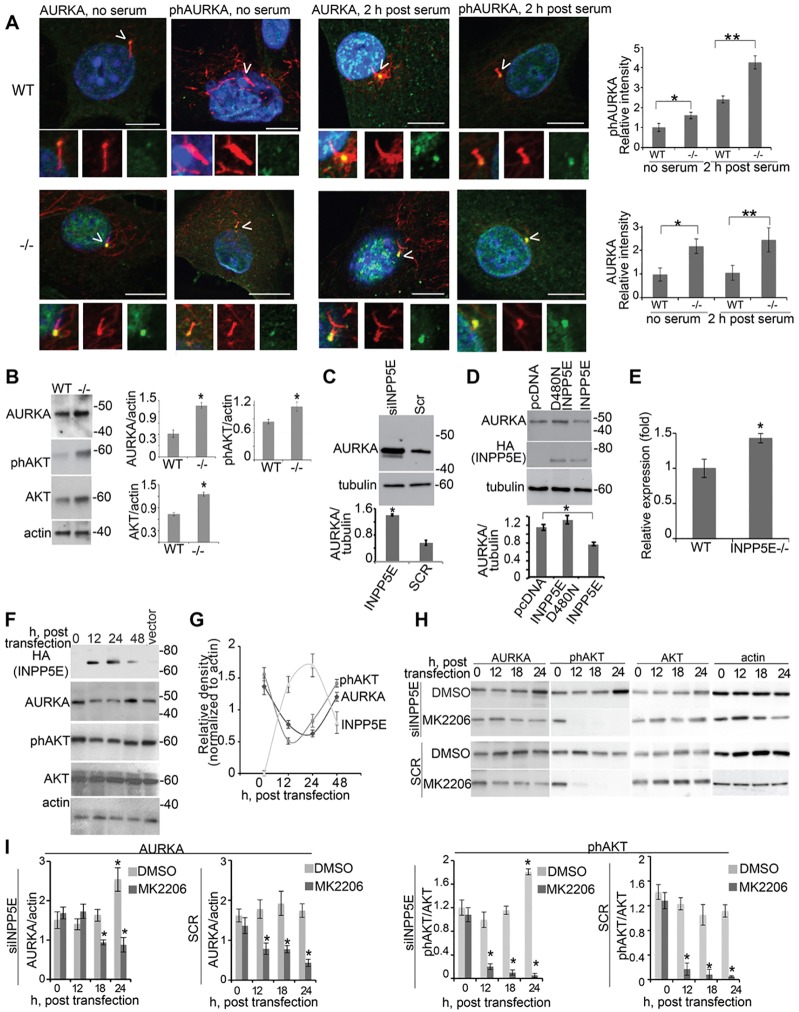
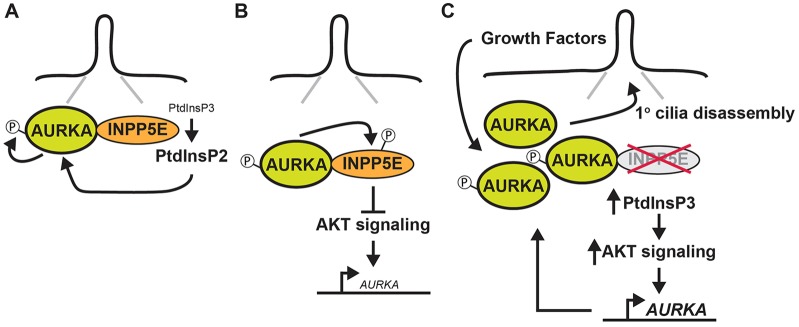
Mutations in inositol polyphosphate 5-phosphatase E (INPP5E) cause the ciliopathies known as Joubert and MORM syndromes; however, the role of INPP5E in ciliary biology is not well understood. Here, we describe an interaction between INPP5E and AURKA, a centrosomal kinase that regulates mitosis and ciliary disassembly, and we show that this interaction is important for the stability of primary cilia. Furthermore, AURKA phosphorylates INPP5E and thereby increases its 5-phosphatase activity, which in turn promotes transcriptional downregulation of AURKA, partly through an AKT-dependent mechanism. These findings establish the first direct link between AURKA and phosphoinositide signaling and suggest that the function of INPP5E in cilia is at least partly mediated by its interactions with AURKA.
Keywords: AURKA; INPP5E; Primary cilium.
© 2015. Published by The Company of Biologists Ltd.
Figures

References
-
- Bielas S. L., Silhavy J. L., Brancati F., Kisseleva M. V., Al-Gazali L., Sztriha L., Bayoumi R. A., Zaki M. S., Abdel-Aleem A., Rosti R. O. <et al.(2009). Mutations in INPP5E, encoding inositol polyphosphate-5-phosphatase E, link phosphatidyl inositol signaling to the ciliopathies. Nat. Genet. 41, 1032–1036 10.1038/ng.423 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
