

FRIZZY PANICLE drives supernumerary spikelets in bread wheat
- PMID: 25398545
- PMCID: PMC4281007
- DOI: 10.1104/pp.114.250043
FRIZZY PANICLE drives supernumerary spikelets in bread wheat
Abstract
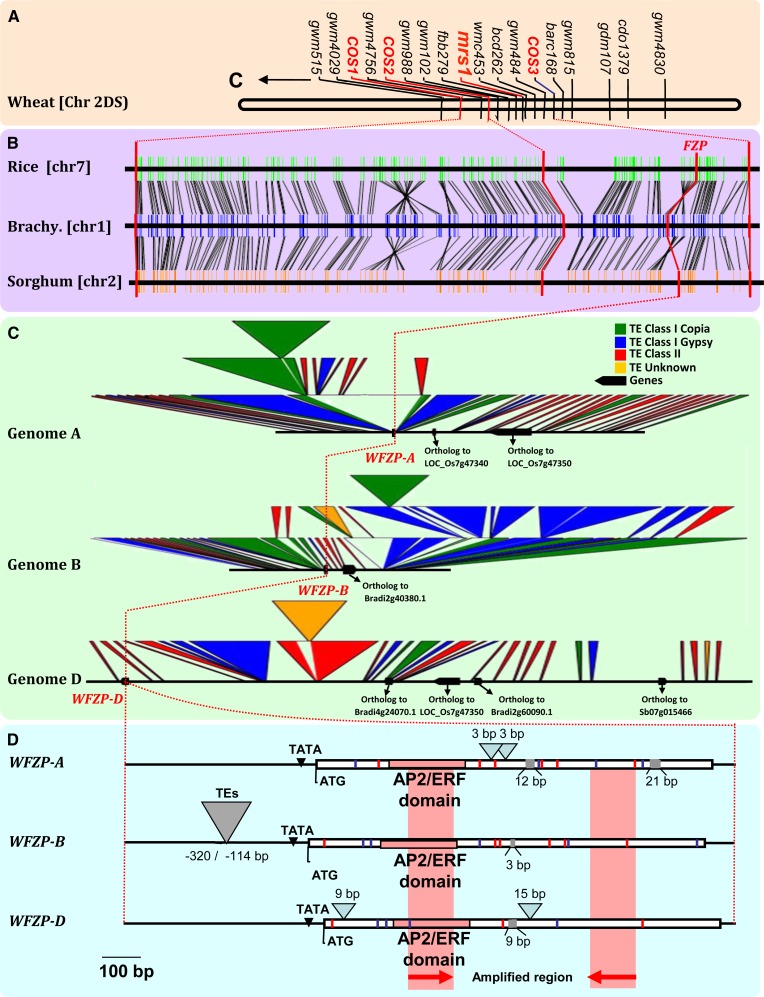
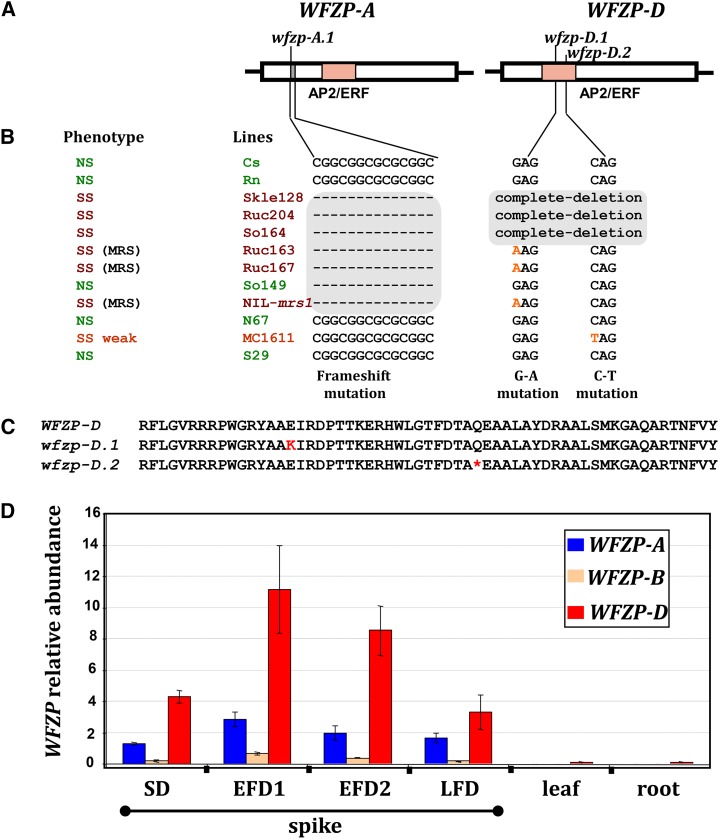
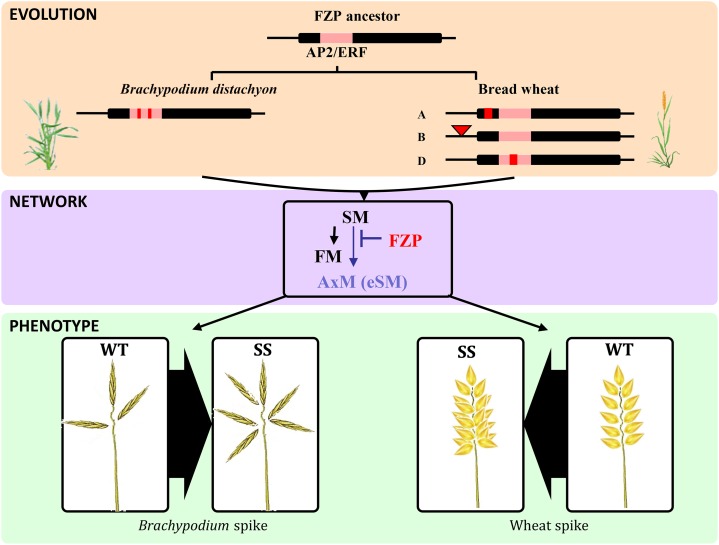
Bread wheat (Triticum aestivum) inflorescences, or spikes, are characteristically unbranched and normally bear one spikelet per rachis node. Wheat mutants on which supernumerary spikelets (SSs) develop are particularly useful resources for work towards understanding the genetic mechanisms underlying wheat inflorescence architecture and, ultimately, yield components. Here, we report the characterization of genetically unrelated mutants leading to the identification of the wheat FRIZZY PANICLE (FZP) gene, encoding a member of the APETALA2/Ethylene Response Factor transcription factor family, which drives the SS trait in bread wheat. Structural and functional characterization of the three wheat FZP homoeologous genes (WFZP) revealed that coding mutations of WFZP-D cause the SS phenotype, with the most severe effect when WFZP-D lesions are combined with a frameshift mutation in WFZP-A. We provide WFZP-based resources that may be useful for genetic manipulations with the aim of improving bread wheat yield by increasing grain number.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures


References
-
- Acevedo E, Silva P, Silva H (2002) Wheat growth and physiology. InCurtis B, Rajaram S, Gomez Macpherson H, eds, Bread Wheat Improvement and Production. Food and Agriculture Organization of the United Nations, Rome
-
- Amagai Y, Martinek P, Watanabe N, Kuboyama T (2014) Microsatellite mapping of genes for branched spike and soft glumes in Triticum monococcum L. Genet Resour Crop Evol 61: 465–471
-
- Chuck G, Muszynski M, Kellogg E, Hake S, Schmidt RJ (2002) The control of spikelet meristem identity by the branched silkless1 gene in maize. Science 298: 1238–1241 - PubMed
-
- Coffman FA. (1924) Supernumerary spikelets in Mindum wheat. J Hered 5: 187–192
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
