

Differential role of STIM1 and STIM2 during transient inward (T in) current generation and the maturation process in the Xenopus oocyte
- PMID: 25399338
- PMCID: PMC4236480
- DOI: 10.1186/s12899-014-0009-x
Differential role of STIM1 and STIM2 during transient inward (T in) current generation and the maturation process in the Xenopus oocyte
Abstract
Background: The Xenopus oocyte is a useful cell model to study Ca2+ homeostasis and cell cycle regulation, two highly interrelated processes. Here, we used antisense oligonucleotides to investigate the role in the oocyte of stromal interaction molecule (STIM) proteins that are fundamental elements of the store-operated calcium-entry (SOCE) phenomenon, as they are both sensors for Ca2+ concentration in the intracellular reservoirs as well as activators of the membrane channels that allow Ca2+ influx.
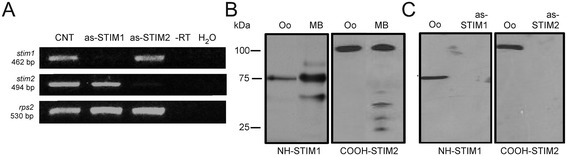
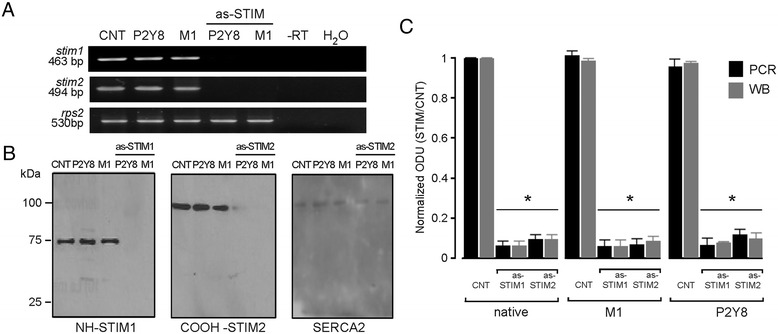
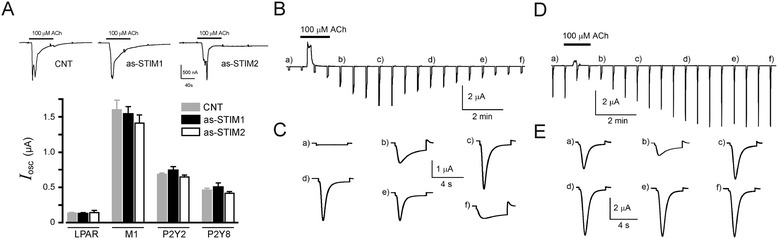
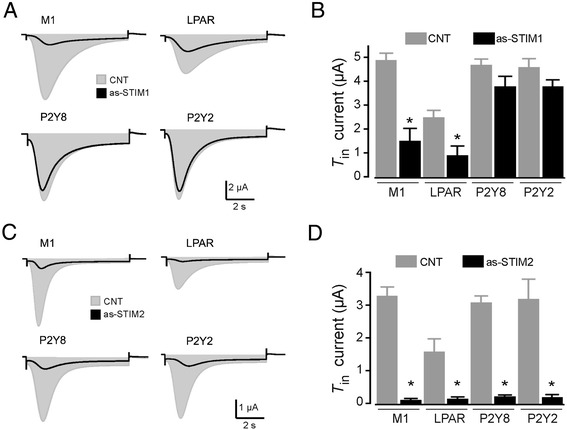
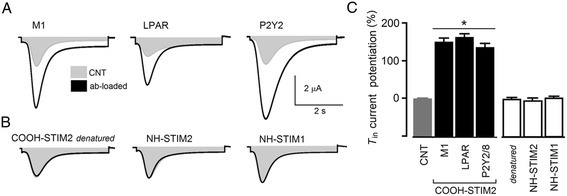
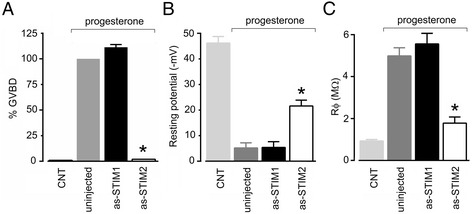
Results: Endogenous STIM1 and STIM2 expression was demonstrated, and their synthesis was knocked down 48-72 h after injecting oocytes with specific antisense sequences. Selective elimination of their mRNA and protein expression was confirmed by PCR and Western blot analysis, and we then evaluated the effect of their absence on two endogenous responses: the opening of SOC channels elicited by G protein-coupled receptor (GPCR)-activated Ca2+ release, and the process of maturation stimulated by progesterone. Activation of SOC channels was monitored electrically by measuring the T in response, a Ca2+-influx-dependent Cl- current, while maturation was assessed by germinal vesicle breakdown (GVBD) scoring and electrophysiology.
Conclusions: It was found that STIM2, but not STIM1, was essential in both responses, and T in currents and GVBD were strongly reduced or eliminated in cells devoid of STIM2; STIM1 knockdown had no effect on the maturation process, but it reduced the T in response by 15 to 70%. Thus, the endogenous SOCE response in Xenopus oocytes depended mainly on STIM2, and its expression was necessary for entry into meiosis induced by progesterone.
Figures
Similar articles
-
Calcium signaling in mouse oocyte maturation: the roles of STIM1, ORAI1 and SOCE.Mol Hum Reprod. 2012 Apr;18(4):194-203. doi: 10.1093/molehr/gar071. Epub 2011 Nov 3. Mol Hum Reprod. 2012. PMID: 22053056
-
STIM1 but not STIM2 is an essential regulator of Ca2+ influx-mediated NADPH oxidase activity in neutrophil-like HL-60 cells.Biochem Pharmacol. 2009 Sep 1;78(5):504-13. doi: 10.1016/j.bcp.2009.05.006. Epub 2009 May 9. Biochem Pharmacol. 2009. PMID: 19433064
-
A key role for STIM1 in store operated calcium channel activation in airway smooth muscle.Respir Res. 2006 Sep 20;7(1):119. doi: 10.1186/1465-9921-7-119. Respir Res. 2006. PMID: 16987424 Free PMC article.
-
[Store-operated Calcium Entry into B Cells Regulates Autoimmune Inflammation].Yakugaku Zasshi. 2016;136(3):473-8. doi: 10.1248/yakushi.15-00246-3. Yakugaku Zasshi. 2016. PMID: 26935089 Review. Japanese.
-
Remodelling of the endoplasmic reticulum during store-operated calcium entry.Biol Cell. 2011 Aug;103(8):365-80. doi: 10.1042/BC20100152. Biol Cell. 2011. PMID: 21736554 Review.
Cited by
-
Molecular physiology and pathophysiology of stromal interaction molecules.Exp Biol Med (Maywood). 2018 Mar;243(5):451-472. doi: 10.1177/1535370218754524. Epub 2018 Jan 24. Exp Biol Med (Maywood). 2018. PMID: 29363328 Free PMC article. Review.
-
Interplay between ER Ca2+ Binding Proteins, STIM1 and STIM2, Is Required for Store-Operated Ca2+ Entry.Int J Mol Sci. 2018 May 19;19(5):1522. doi: 10.3390/ijms19051522. Int J Mol Sci. 2018. PMID: 29783744 Free PMC article.
-
Identification of a unique endoplasmic retention motif in the Xenopus GIRK5 channel and its contribution to oocyte maturation.FEBS Open Bio. 2021 Apr;11(4):1093-1108. doi: 10.1002/2211-5463.13113. Epub 2021 Mar 3. FEBS Open Bio. 2021. PMID: 33565726 Free PMC article.
References
-
- Parekh AB, Penner R. Store depletion and calcium influx. Physiol Rev. 1997;77:901–930. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
