

Review
doi: 10.1016/B978-0-12-397918-6.00001-X.
Catalytic mechanisms and regulation of protein kinases
Affiliations
- PMID: 25399640
- PMCID: PMC4373616
- DOI: 10.1016/B978-0-12-397918-6.00001-X
Item in Clipboard
Review
Catalytic mechanisms and regulation of protein kinases
Methods Enzymol.
2014.
Abstract
Protein kinases transfer a phosphoryl group from ATP onto target proteins and play a critical role in signal transduction and other cellular processes. Here, we review the kinase kinetic and chemical mechanisms and their application in understanding kinase structure and function. Aberrant kinase activity has been implicated in many human diseases, in particular cancer. We highlight applications of technologies and concepts derived from kinase mechanistic studies that have helped illuminate how kinases are regulated and contribute to pathophysiology.
Keywords: B-Raf; Bisubstrate analog; Chemical rescue; EGFR; Erlotinib; Inhibitor; Lapatinib; Src; Transition state.
Figures
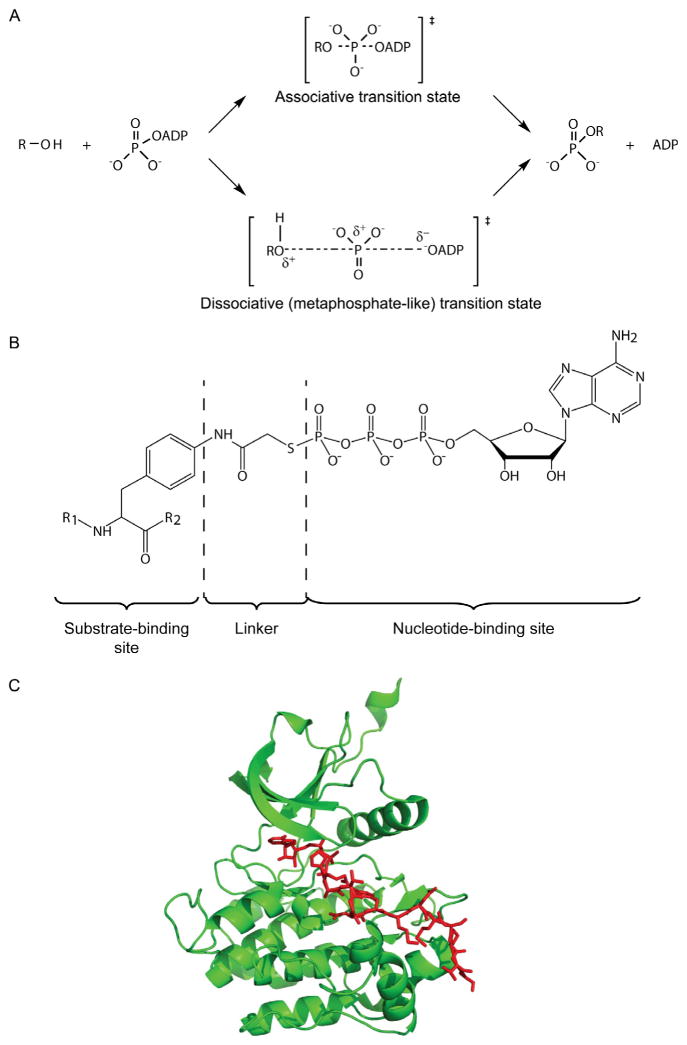
Protein kinase mechanism and bisubstrate analog inhibitors. (A) Mechanistic scheme of associative and dissociative transition states of phosphoryl transfer. A dissociative state model is proposed for the transfer of the γ-phosphoryl group of ATP to hydroxy group in a kinase–substrate. (B) Designed bisubstrate analog inhibitor for a protein tyrosine kinase. R1 and R2 amino acid sequences are derived from efficient substrate motifs. The linker is predicted to result in a 5–5.7 Å spacer between the substrate-binding site and the nucleotide-binding site, compatible with a dissociative transition state. ATPγS was used as the ATP-mimic analog. (C) The crystal structure of IRK (green ribbon) in complex with the above peptide–ATP conjugate (red stick), determined at 2.7 Å resolution (PDB code: 1GAG; Parang et al., 2001).
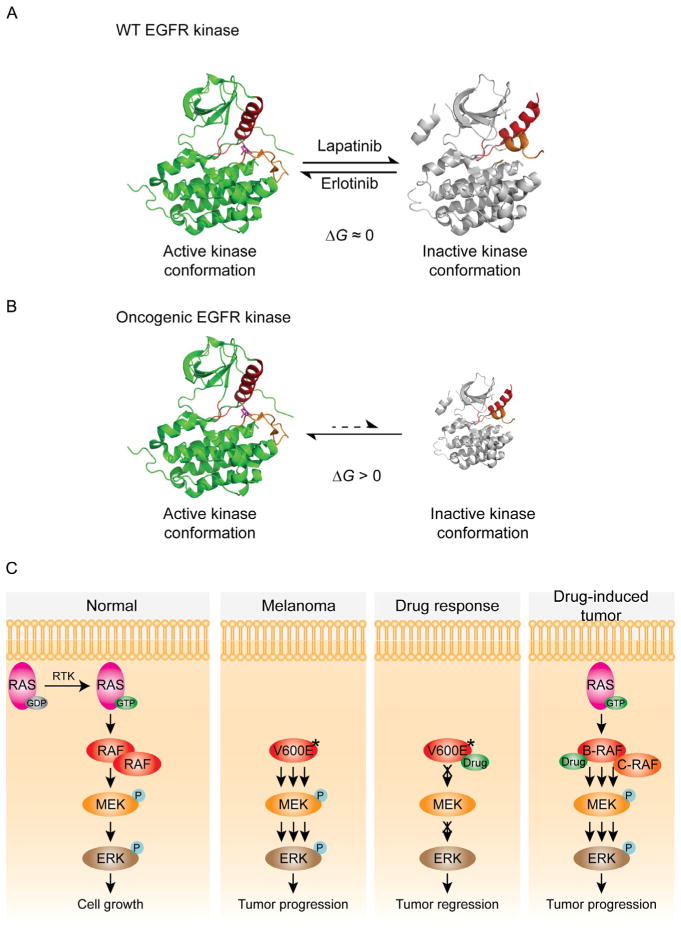
Oncogenic kinase mutant mechanisms. (A) The crystal structures of active and inactive EGFR kinase conformations are shown in ribbon representation. Two key structural elements within the kinase domain, the activation loop and helix, are colored orange and red. For WT EGF-bound EGFR, the active kinase conformation (green ribbon, PDB code: 1M17) (Stamos et al., 2002) is nearly isoenergetic to the inactive kinase conformation (gray ribbon, PDB code: 1XKK). Erlotinib selectively binds and stabilizes the active conformation; therefore, the active conformation is dominant in the presence of erlotinib. Similarly, the inactive conformation is dominant in the presence of lapatinib (Wood et al., 2004). Erlotinib and lapatinib are both potent inhibitors against WT EGFR. (B) Oncogenic mutations alter the energy landscape to favor the active conformation. As a result, lapatinib has little access to the majority of oncogenic EGFR mutants, which are locked into the active conformation. The size of each structure has been schematized to depict the relative population of active and inactive conformations. (C) In normal cells, activated RAS recruits B-RAF from the cytosol to the plasma membrane for dimerization and activation. Once activated, B-RAF phosphorylates MEK, which in turn phosphorylates and activates ERK. Oncogenic forms of B-RAF (marked with V600E*) highly activate the ERK pathway in a RAS-independent manner. In B-RAF mutant cancer cells, such as melanoma, B-RAF inhibitor efficiently blocks ERK activation. In RAS mutant cancer cells, the same ATP-competitive inhibitor unexpectedly activates the ERK pathway by promoting RAF dimerization.
References
-
- Ablooglu AJ, Frankel M, Rusinova E, Ross JB, Kohanski RA. Multiple activation loop conformations and their regulatory properties in the insulin receptor’s kinase domain. Journal of Biological Chemistry. 2001;276:46933–46940. - PubMed
-
- Ablooglu AJ, Till JH, Kim K, Parang K, Cole PA, Hubbard SR, et al. Probing the catalytic mechanism of the insulin receptor kinase with a tetrafluorotyrosine-containing peptide substrate. Journal of Biological Chemistry. 2000;275:30394–30398. - PubMed
-
- Adams JA. Kinetic and catalytic mechanisms of protein kinases. Chemical Reviews. 2001;101:2271–2290. - PubMed
-
- Adams JA, Taylor SS. Energetic limits of phosphotransfer in the catalytic subunit of cAMP-dependent protein kinase as measured by viscosity experiments. Biochemistry. 1992;31:8516–8522. - PubMed
-
- Admiraal SJ, Herschlag D. Mapping the transition state for ATP hydrolysis: Implications for enzymatic catalysis. Chemistry & Biology. 1995;2:729–739. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
