

Brain composition in Godyris zavaleta, a diurnal butterfly, Reflects an increased reliance on olfactory information
- PMID: 25400217
- PMCID: PMC4354442
- DOI: 10.1002/cne.23711
Brain composition in Godyris zavaleta, a diurnal butterfly, Reflects an increased reliance on olfactory information
Abstract
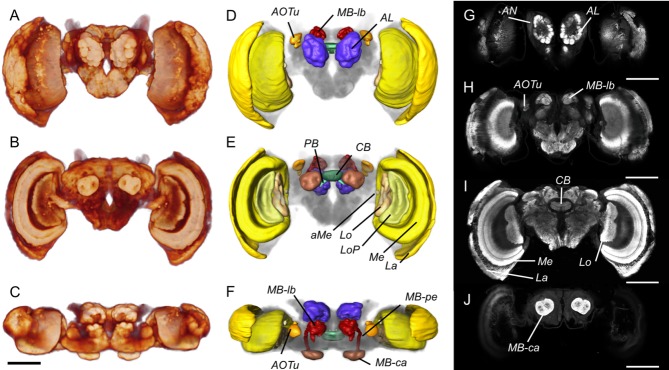
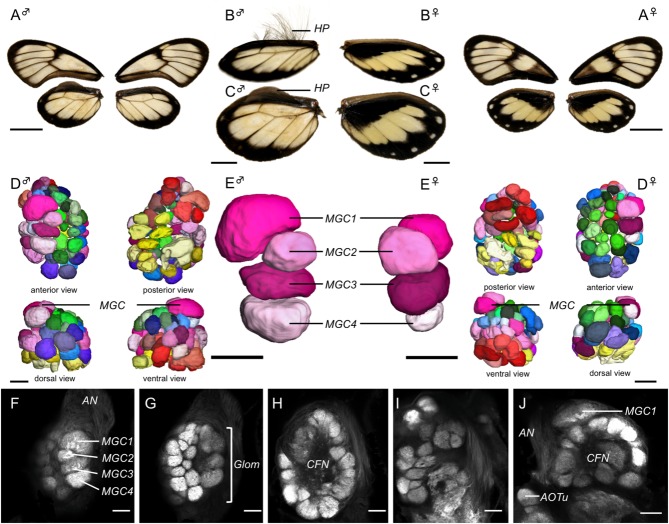
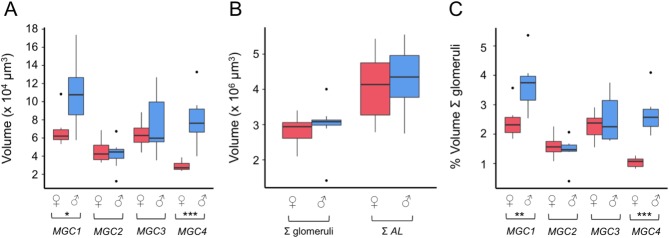
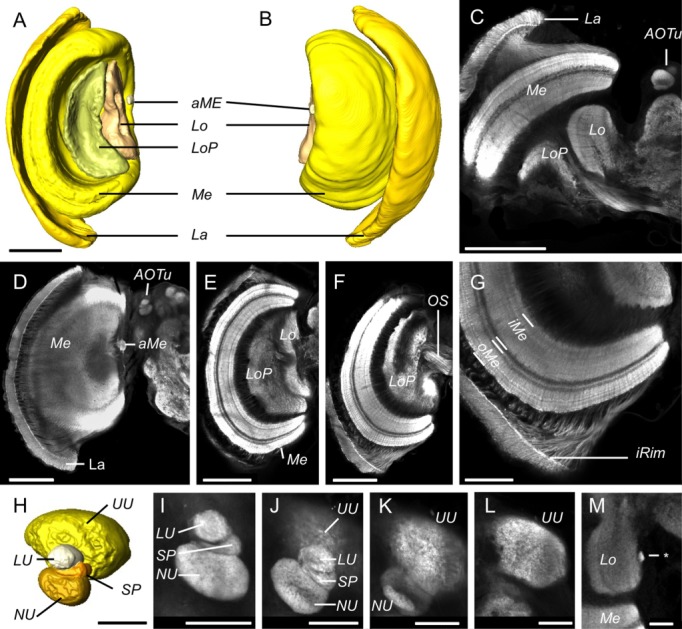
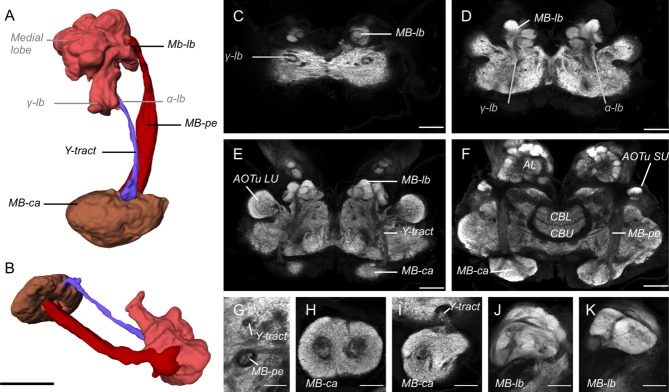
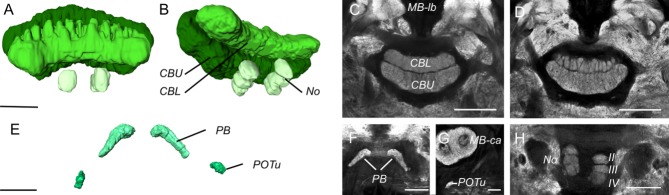
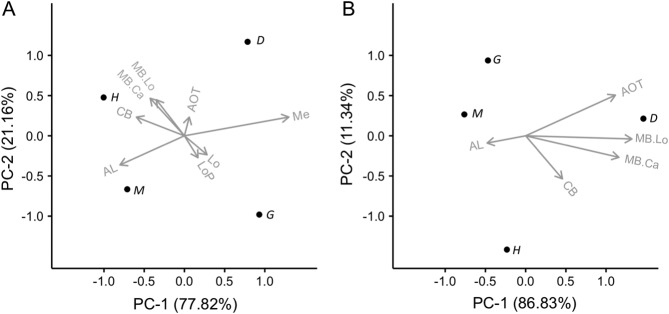
Interspecific comparisons of brain structure can inform our functional understanding of brain regions, identify adaptations to species-specific ecologies, and explore what constrains adaptive changes in brain structure, and coevolution between functionally related structures. The value of such comparisons is enhanced when the species considered have known ecological differences. The Lepidoptera have long been a favored model in evolutionary biology, but to date descriptions of brain anatomy have largely focused on a few commonly used neurobiological model species. We describe the brain of Godyris zavaleta (Ithomiinae), a member of a subfamily of Neotropical butterflies with enhanced reliance on olfactory information. We demonstrate for the first time the presence of sexually dimorphic glomeruli within a distinct macroglomerular complex (MGC) in the antennal lobe of a diurnal butterfly. This presents a striking convergence with the well-known moth MGC, prompting a discussion of the potential mechanisms behind the independent evolution of specialized glomeruli. Interspecific analyses across four Lepidoptera further show that the relative size of sensory neuropils closely mirror interspecific variation in sensory ecology, with G. zavaleta displaying levels of sensory investment intermediate between the diurnal monarch butterfly (Danaus plexippus), which invests heavily in visual neuropil, and night-flying moths, which invest more in olfactory neuropil. We identify several traits that distinguish butterflies from moths, and several that distinguish D. plexippus and G. zavaleta. Our results illustrate that ecological selection pressures mold the structure of invertebrate brains, and exemplify how comparative analyses across ecologically divergent species can illuminate the functional significance of variation in brain structure.
Keywords: Godyris zavaleta; Ithomiinae; Lepidoptera; adaptive brain evolution; antennal lobe; comparative neuroanatomy; macroglomerular complex; sexual dimorphism.
© 2014 Wiley Periodicals, Inc.
Figures
References
-
- Ali FA. Structure and metamorphosis of the brain and suboesophageal ganglion of Pieris brassicae (L.) (Lepidoptera: Pieridae) Trans R Entomol Soc Lond. 1974;125:363–412.
-
- Andersson S, Dobson HEM. Behavioral foraging responses by the butterfly Heliconius melpomene to Lantana camara floral scent. J Chem Ecol. 2003;29:2303–2318. - PubMed
-
- Andersson J, Borg-Karlson A-K, Vongvanich N, Wiklund C. Male sex pheromone release and female mate choice in a butterfly. J Exp Biol. 2007;210:964–970. - PubMed
-
- Barton RA, Harvey PH. Mosaic evolution of brain structure in mammals. Nature. 2000;405:1055–1058. - PubMed
-
- Barton RA, Purvis A, Harvey PH. Evolutionary radiation of visual and olfactory brain systems in primates, bats and insectivores. Philos Trans R Soc Lond B Biol Sci. 1995;348:381–392. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
