

RNA cytidine acetyltransferase of small-subunit ribosomal RNA: identification of acetylation sites and the responsible acetyltransferase in fission yeast, Schizosaccharomyces pombe
- PMID: 25402480
- PMCID: PMC4234376
- DOI: 10.1371/journal.pone.0112156
RNA cytidine acetyltransferase of small-subunit ribosomal RNA: identification of acetylation sites and the responsible acetyltransferase in fission yeast, Schizosaccharomyces pombe
Abstract
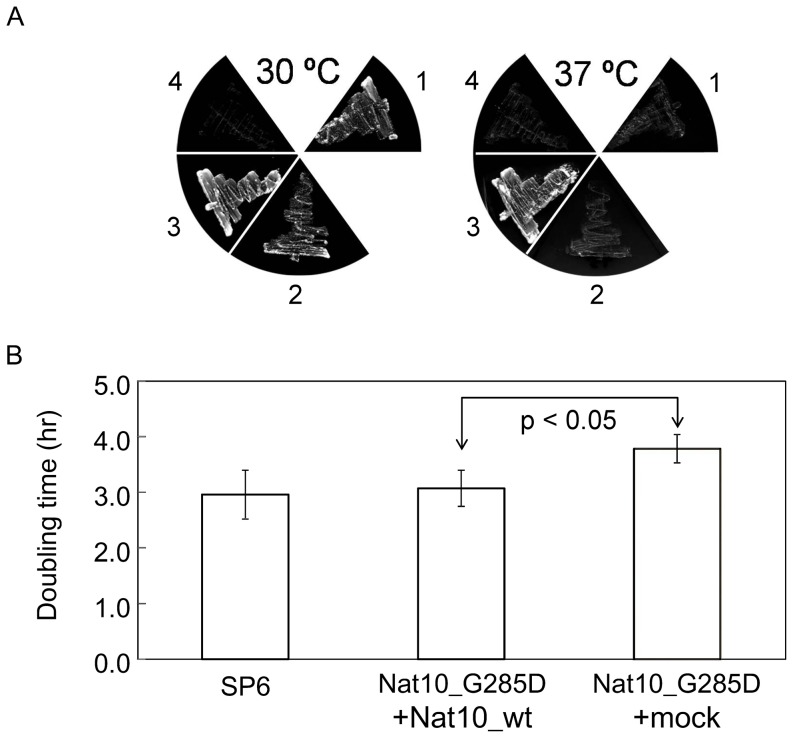
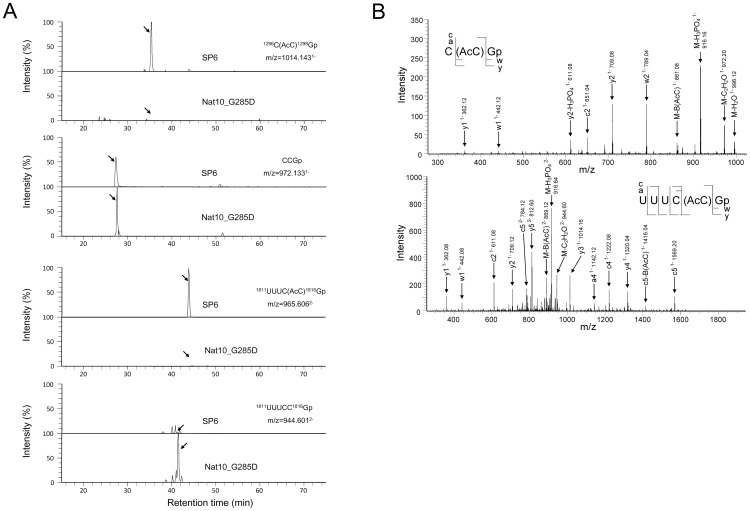
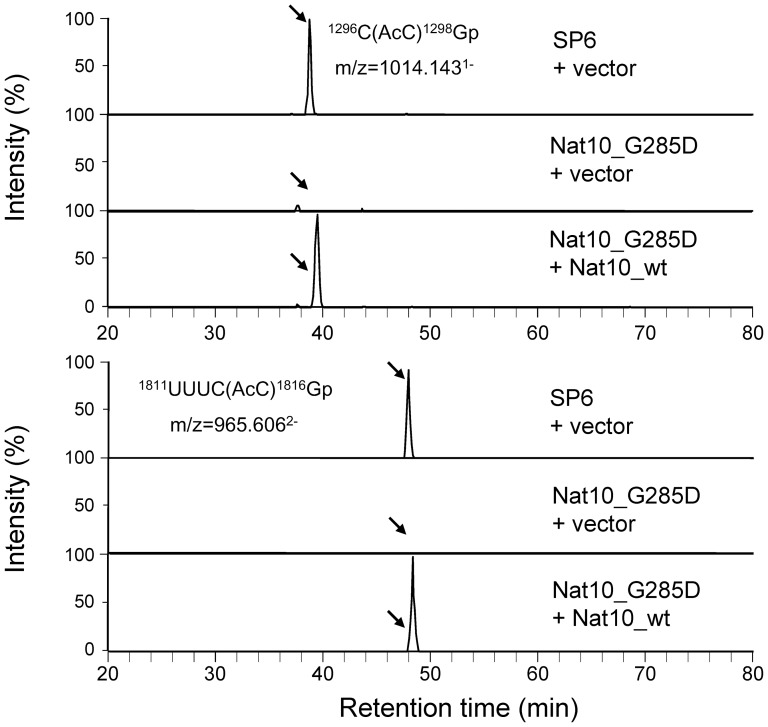
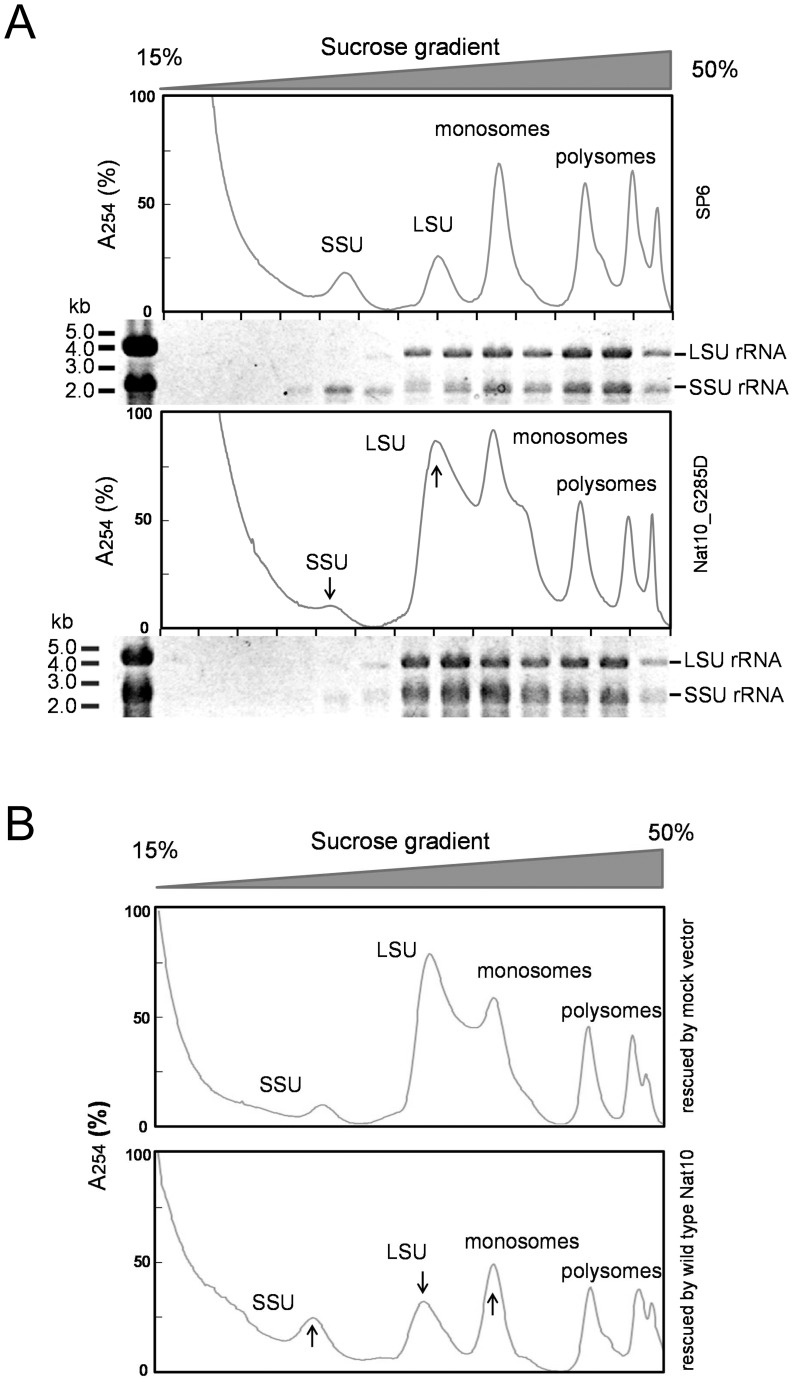
The eukaryotic small-subunit (SSU) ribosomal RNA (rRNA) has two evolutionarily conserved acetylcytidines. However, the acetylation sites and the acetyltransferase responsible for the acetylation have not been identified. We performed a comprehensive MS-based analysis covering the entire sequence of the fission yeast, Schizosaccharomyces pombe, SSU rRNA and identified two acetylcytidines at positions 1297 and 1815 in the 3' half of the rRNA. To identify the enzyme responsible for the cytidine acetylation, we searched for an S. pombe gene homologous to TmcA, a bacterial tRNA N-acetyltransferase, and found one potential candidate, Nat10. A temperature-sensitive strain of Nat10 with a mutation in the Walker A type ATP-binding motif abolished the cytidine acetylation in SSU rRNA, and the wild-type Nat10 supplemented to this strain recovered the acetylation, providing evidence that Nat10 is necessary for acetylation of SSU rRNA. The Nat10 mutant strain showed a slow-growth phenotype and was defective in forming the SSU rRNA from the precursor RNA, suggesting that cytidine acetylation is necessary for ribosome assembly.
Conflict of interest statement
Figures
References
-
- Yusupova G, Yusupov M (2014) High-Resolution Structure of the Eukaryotic 80S Ribosome. Annu Rev Biochem 83: 467–486. - PubMed
-
- Voigts-Hoffmann F, Klinge S, Ban N (2012) Structural insights into eukaryotic ribosomes and the initiation of translation. Curr Opin Struct Biol 22: 768–777. - PubMed
-
- Jenner L, Melnikov S, Garreau de Loubresse N, Ben-Shem A, Iskakova M, et al. (2012) Crystal structure of the 80S yeast ribosome. Curr Opin Struct Biol 22: 759–767. - PubMed
-
- Klinge S, Voigts-Hoffmann F, Leibundgut M, Ban N (2012) Atomic structures of the eukaryotic ribosome. Trends Biochem Sci 37: 189–198. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
