

Telomere position effect: regulation of gene expression with progressive telomere shortening over long distances
- PMID: 25403178
- PMCID: PMC4233240
- DOI: 10.1101/gad.251041.114
Telomere position effect: regulation of gene expression with progressive telomere shortening over long distances
Abstract
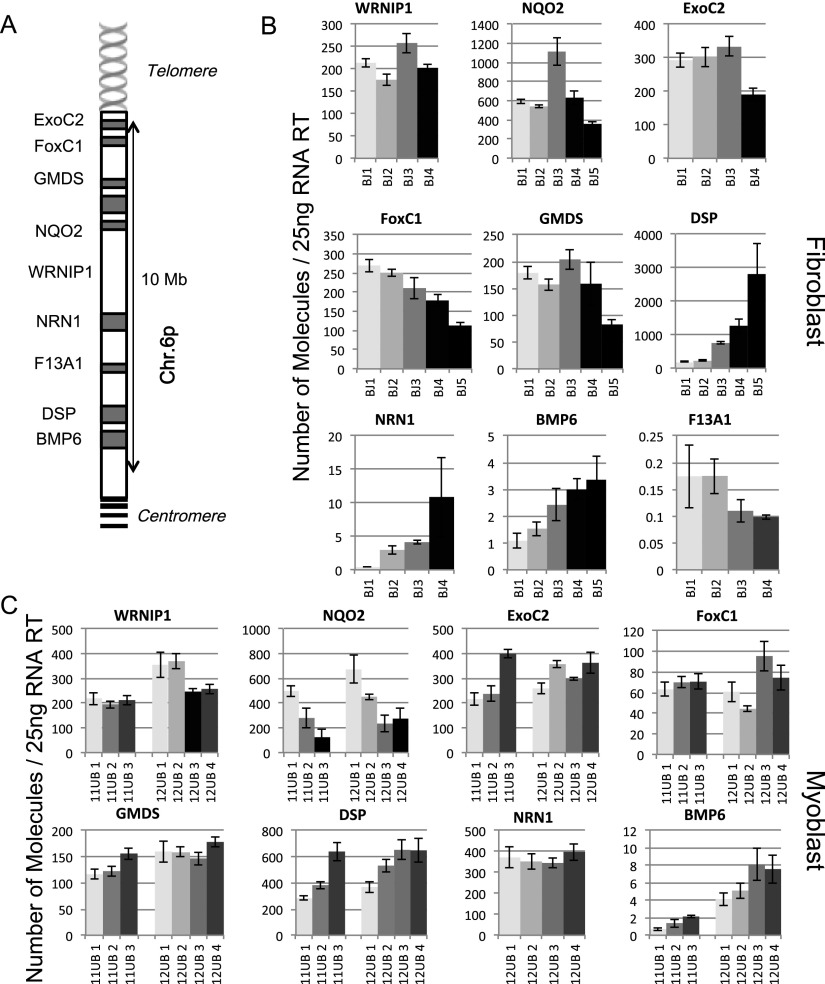
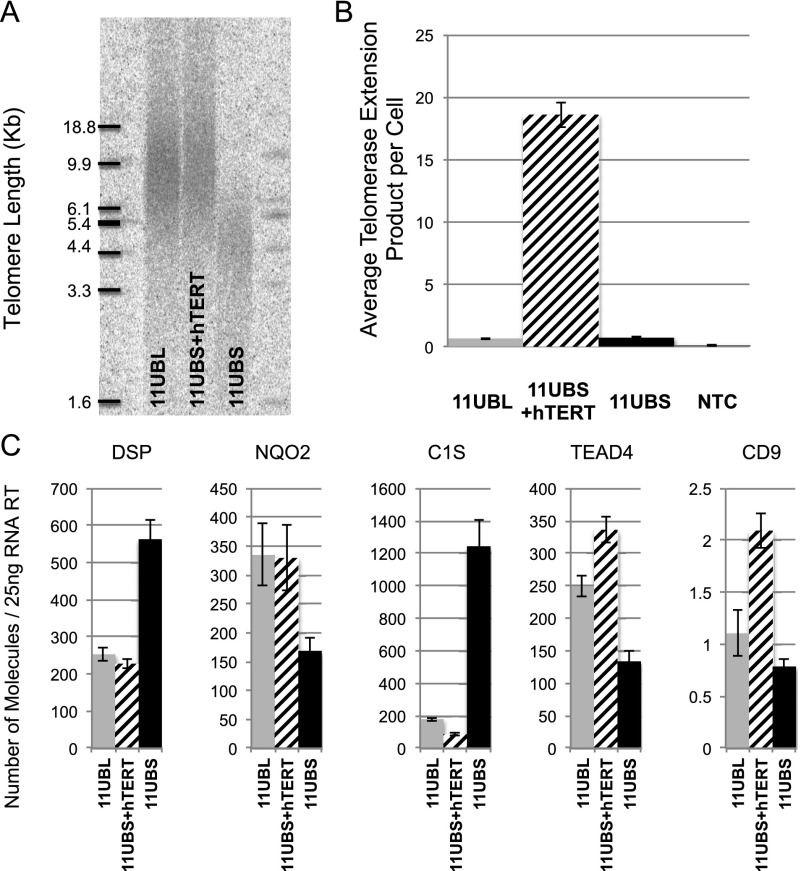
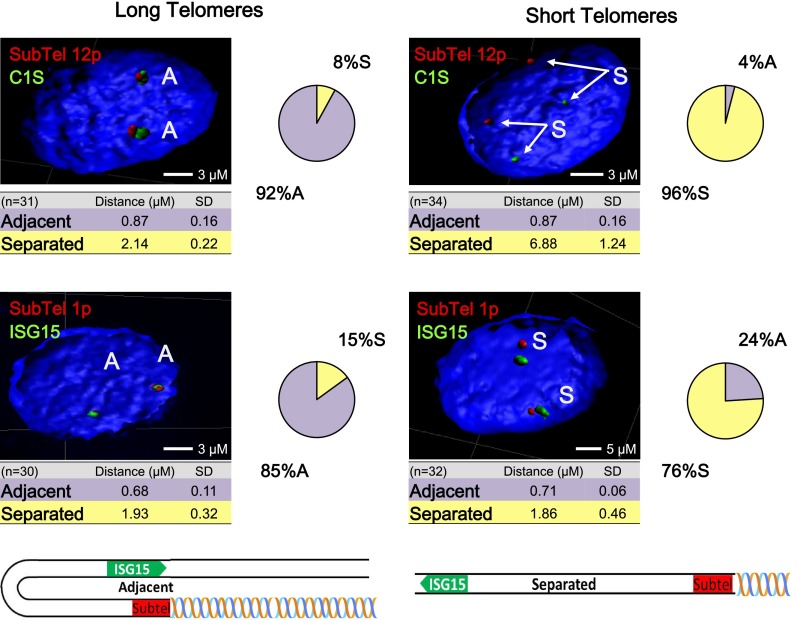
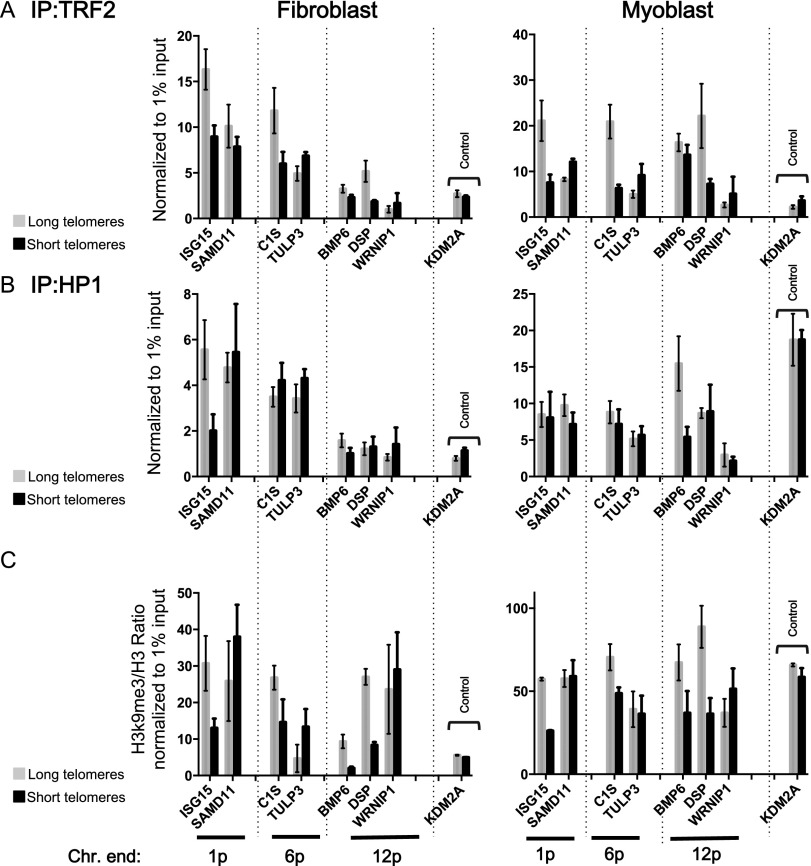
While global chromatin conformation studies are emerging, very little is known about the chromatin conformation of human telomeres. Most studies have focused on the role of telomeres as a tumor suppressor mechanism. Here we describe how telomere length regulates gene expression long before telomeres become short enough to produce a DNA damage response (senescence). We directly mapped the interactions adjacent to specific telomere ends using a Hi-C (chromosome capture followed by high-throughput sequencing) technique modified to enrich for specific genomic regions. We demonstrate that chromosome looping brings the telomere close to genes up to 10 Mb away from the telomere when telomeres are long and that the same loci become separated when telomeres are short. Furthermore, expression array analysis reveals that many loci, including noncoding RNAs, may be regulated by telomere length. We report three genes (ISG15 [interferon-stimulated gene 15 kd], DSP [Desmoplakin], and C1S [complement component 1s subcomplement]) located at three different subtelomeric ends (1p, 6p, and 12p) whose expressions are altered with telomere length. Additionally, we confirmed by in situ analysis (3D-FISH [three-dimensional fluorescence in situ hybridization]) that chromosomal looping occurs between the loci of those genes and their respective telomere ends. We term this process TPE-OLD for "telomere position effect over long distances." Our results suggest a potential novel mechanism for how telomere shortening could contribute to aging and disease initiation/progression in human cells long before the induction of a critical DNA damage response.
Keywords: age-dependent gene expression; cancer; chromatin; chromosome looping; replicative aging; senescence; telomerase.
© 2014 Robin et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
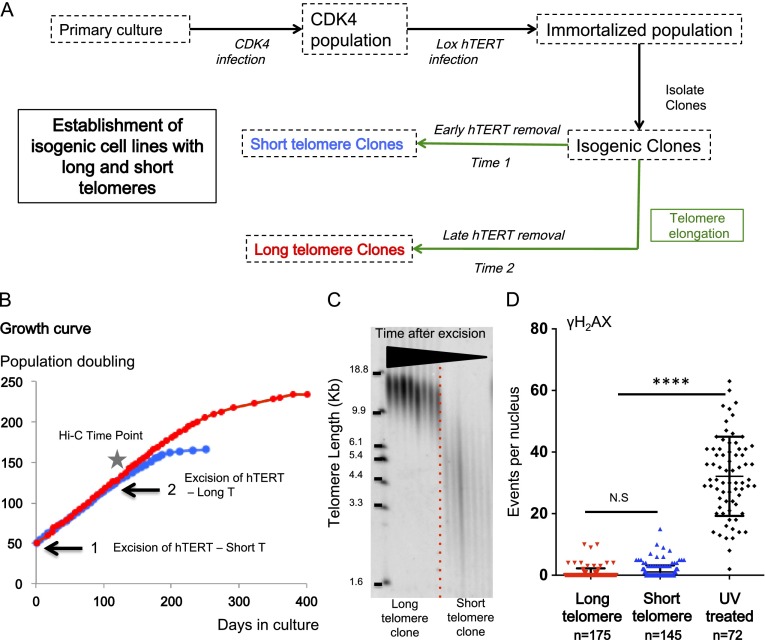
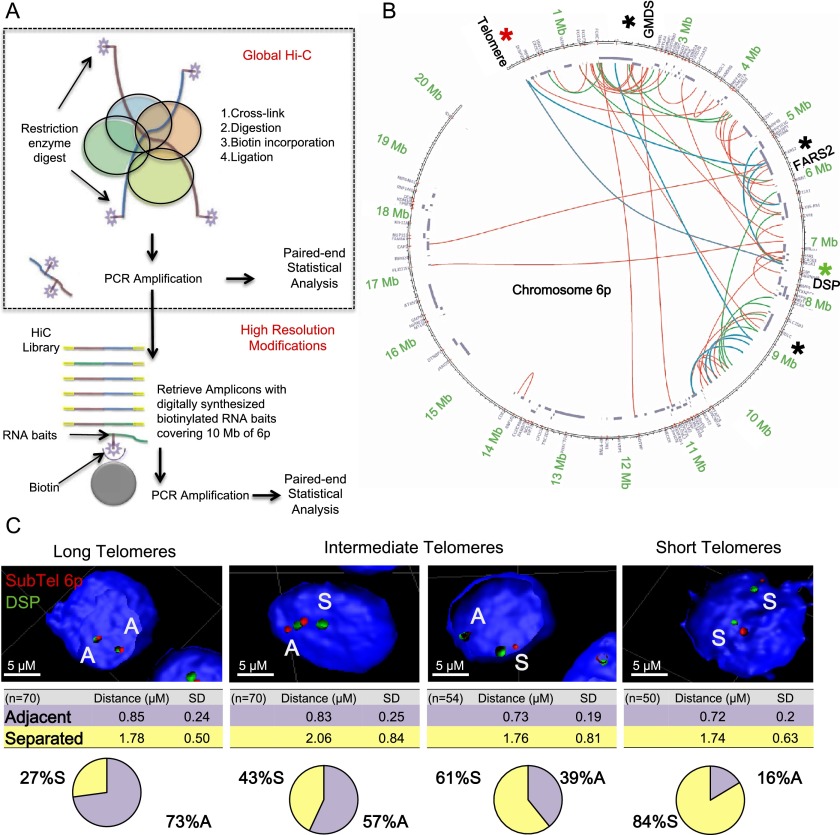
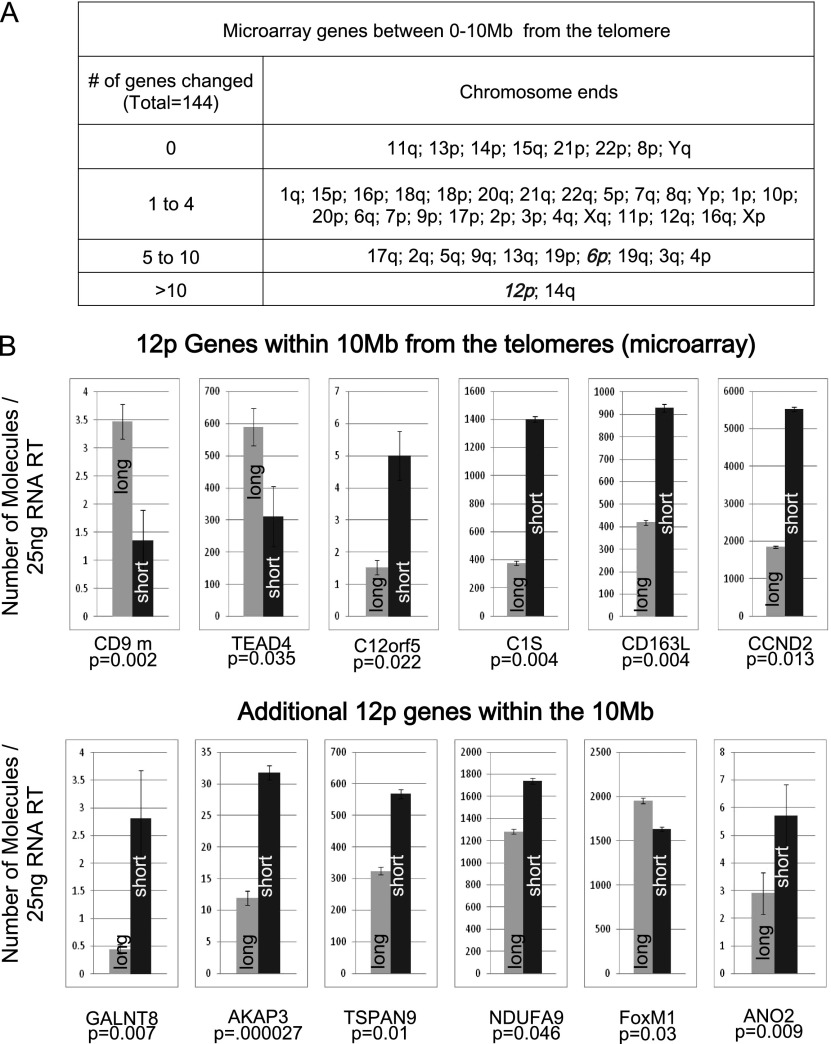
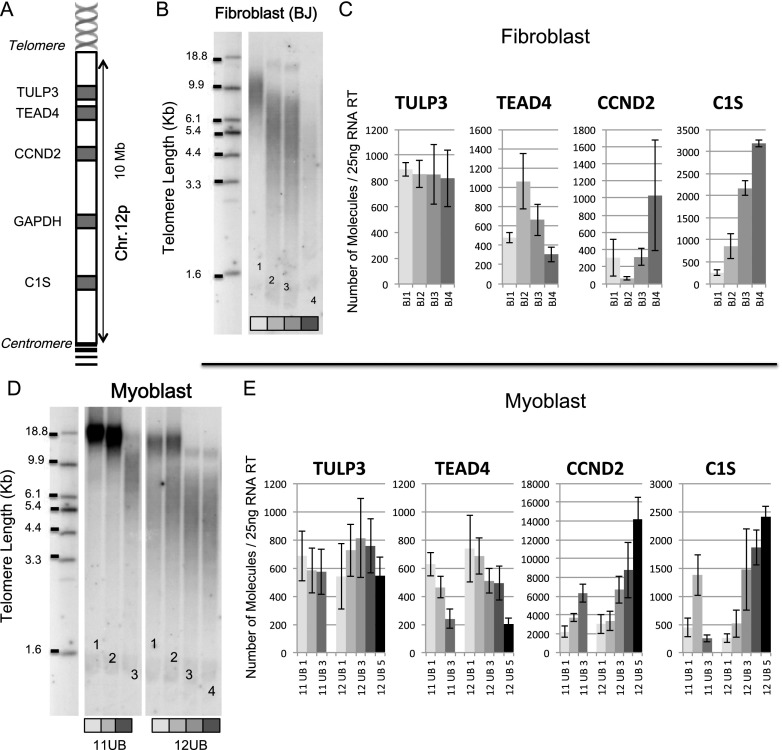
Comment in
-
The long reach of telomeres.Genes Dev. 2014 Nov 15;28(22):2445-6. doi: 10.1101/gad.254573.114. Genes Dev. 2014. PMID: 25403176 Free PMC article.
-
Chromosome biology: Short telomeres can't reach.Nat Rev Mol Cell Biol. 2014 Dec;15(12):766-7. doi: 10.1038/nrm3914. Nat Rev Mol Cell Biol. 2014. PMID: 25415503 No abstract available.
References
-
- Apostolou E, Thanos D. 2008. Virus infection induces NF-κB-dependent interchromosomal associations mediating monoallelic IFN-β gene expression. Cell 134: 85–96. - PubMed
-
- Arnoult N, Schluth-Bolard C, Letessier A, Drascovic I, Bouarich-Bourimi R, Campisi J, Kim SH, Boussouar A, Ottaviani A, Magdinier F, et al. 2010. Replication timing of human telomeres is chromosome arm-specific, influenced by subtelomeric structures and connected to nuclear localization. PLoS Genet 6: e1000920. - PMC - PubMed
-
- Azzalin CM, Reichenbach P, Khoriauli L, Giulotto E, Lingner J. 2007. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 318: 798–801. - PubMed
-
- Barbieri M, Paolisso G, Kimura M, Gardner JP, Boccardi V, Papa M, Hjelmborg JV, Christensen K, Brimacombe M, Nawrot TS, et al. 2009. Higher circulating levels of IGF-1 are associated with longer leukocyte telomere length in healthy subjects. Mech Ageing Dev 130: 771–776. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous