

Noise decomposition of intracellular biochemical signaling networks using nonequivalent reporters
- PMID: 25404303
- PMCID: PMC4260601
- DOI: 10.1073/pnas.1411932111
Noise decomposition of intracellular biochemical signaling networks using nonequivalent reporters
Abstract
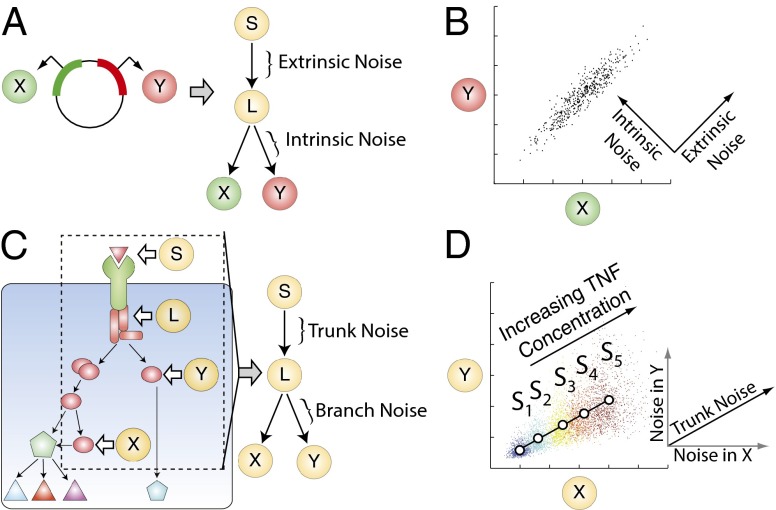
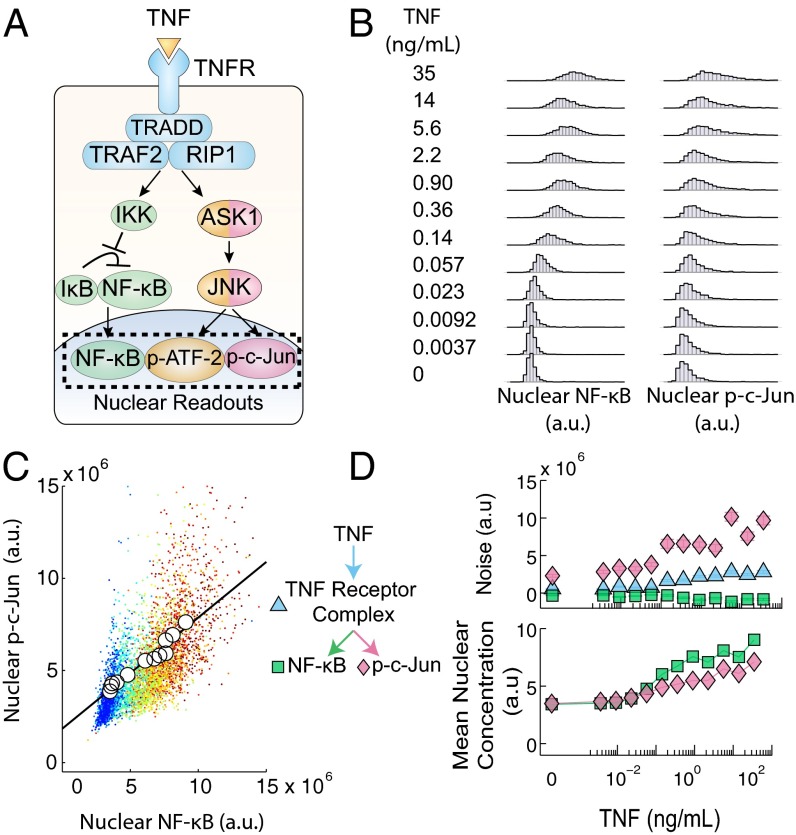
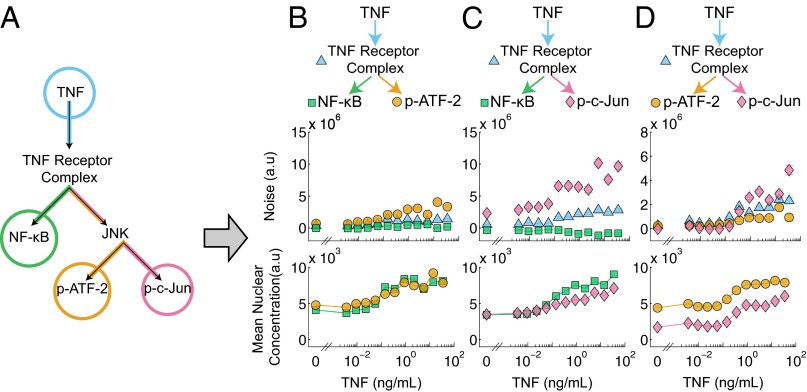
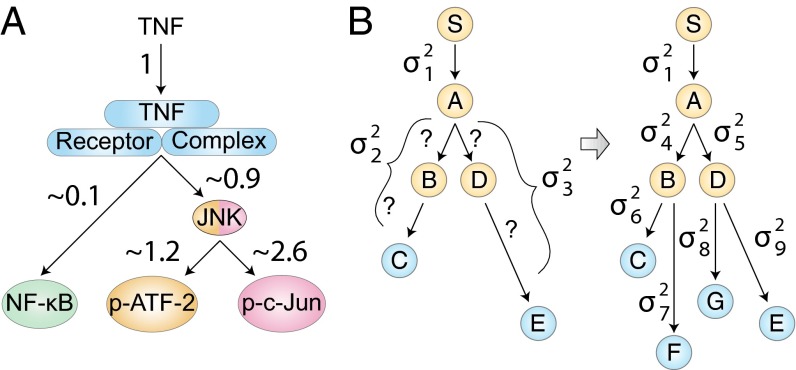
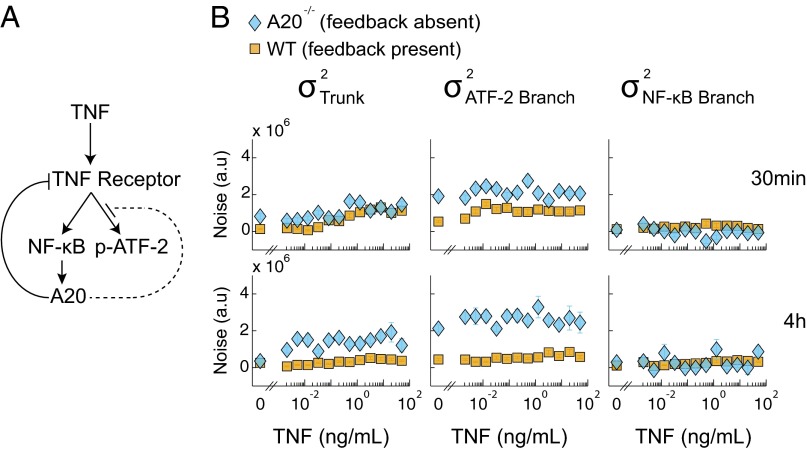
Experimental measurements of biochemical noise have primarily focused on sources of noise at the gene expression level due to limitations of existing noise decomposition techniques. Here, we introduce a mathematical framework that extends classical extrinsic-intrinsic noise analysis and enables mapping of noise within upstream signaling networks free of such restrictions. The framework applies to systems for which the responses of interest are linearly correlated on average, although the framework can be easily generalized to the nonlinear case. Interestingly, despite the high degree of complexity and nonlinearity of most mammalian signaling networks, three distinct tumor necrosis factor (TNF) signaling network branches displayed linearly correlated responses, in both wild-type and perturbed versions of the network, across multiple orders of magnitude of ligand concentration. Using the noise mapping analysis, we find that the c-Jun N-terminal kinase (JNK) pathway generates higher noise than the NF-κB pathway, whereas the activation of c-Jun adds a greater amount of noise than the activation of ATF-2. In addition, we find that the A20 protein can suppress noise in the activation of ATF-2 by separately inhibiting the TNF receptor complex and JNK pathway through a negative feedback mechanism. These results, easily scalable to larger and more complex networks, pave the way toward assessing how noise propagates through cellular signaling pathways and create a foundation on which we can further investigate the relationship between signaling system architecture and biological noise.
Keywords: analysis of variance; extrinsic noise; intrinsic noise; noise decomposition; signal transduction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Information transduction capacity of noisy biochemical signaling networks.Science. 2011 Oct 21;334(6054):354-8. doi: 10.1126/science.1204553. Epub 2011 Sep 15. Science. 2011. PMID: 21921160 Free PMC article.
-
A20 Inhibits β-Cell Apoptosis by Multiple Mechanisms and Predicts Residual β-Cell Function in Type 1 Diabetes.Mol Endocrinol. 2016 Jan;30(1):48-61. doi: 10.1210/me.2015-1176. Epub 2015 Dec 10. Mol Endocrinol. 2016. PMID: 26652732 Free PMC article.
-
TAK1, but not TAB1 or TAB2, plays an essential role in multiple signaling pathways in vivo.Genes Dev. 2005 Nov 15;19(22):2668-81. doi: 10.1101/gad.1360605. Epub 2005 Oct 31. Genes Dev. 2005. PMID: 16260493 Free PMC article.
-
Linking JNK signaling to NF-kappaB: a key to survival.J Cell Sci. 2004 Oct 15;117(Pt 22):5197-208. doi: 10.1242/jcs.01483. J Cell Sci. 2004. PMID: 15483317 Review.
-
[The change in 3-dimensional intracellular parameters alters oscillation pattern of nuclear NF-kappaB].Seikagaku. 2013 Jun;85(6):430-7. Seikagaku. 2013. PMID: 23875470 Review. Japanese. No abstract available.
Cited by
-
Understanding the physical mechanism of intrinsic noise inside viscous isotropic solids.Sci Rep. 2022 Sep 23;12(1):15878. doi: 10.1038/s41598-022-20228-1. Sci Rep. 2022. PMID: 36151467 Free PMC article.
-
Distinct cellular states determine calcium signaling response.Mol Syst Biol. 2016 Dec 15;12(12):894. doi: 10.15252/msb.20167137. Mol Syst Biol. 2016. PMID: 27979909 Free PMC article.
-
Insights about collective decision-making at the genetic level.Biophys Rev. 2020 Feb;12(1):19-24. doi: 10.1007/s12551-019-00608-0. Epub 2019 Dec 16. Biophys Rev. 2020. PMID: 31845181 Free PMC article.
-
Redefining Signaling Pathways with an Expanding Single-Cell Toolbox.Trends Biotechnol. 2016 Jun;34(6):458-469. doi: 10.1016/j.tibtech.2016.02.009. Epub 2016 Mar 9. Trends Biotechnol. 2016. PMID: 26968612 Free PMC article. Review.
-
On Information Extraction and Decoding Mechanisms Improved by Noisy Amplification in Signaling Pathways.Sci Rep. 2019 Oct 7;9(1):14365. doi: 10.1038/s41598-019-50631-0. Sci Rep. 2019. PMID: 31591406 Free PMC article.
References
-
- Dyson S, Gurdon JB. The interpretation of position in a morphogen gradient as revealed by occupancy of activin receptors. Cell. 1998;93(4):557–568. - PubMed
-
- Bronnikov GE, Zhang SJ, Cannon B, Nedergaard J. A dual component analysis explains the distinctive kinetics of cAMP accumulation in brown adipocytes. J Biol Chem. 1999;274(53):37770–37780. - PubMed
-
- McCullagh E, et al. Not all quiet on the noise front. Nat Chem Biol. 2009;5(10):699–704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
