

Relating influenza virus membrane fusion kinetics to stoichiometry of neutralizing antibodies at the single-particle level
- PMID: 25404330
- PMCID: PMC4260548
- DOI: 10.1073/pnas.1411755111
Relating influenza virus membrane fusion kinetics to stoichiometry of neutralizing antibodies at the single-particle level
Abstract
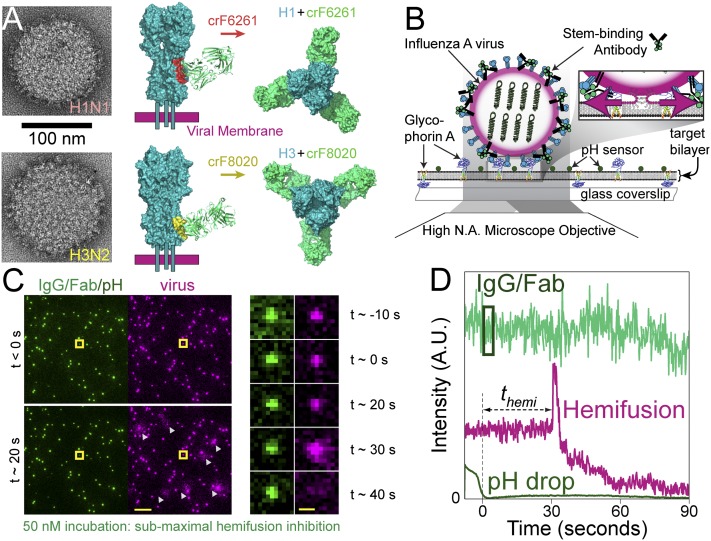
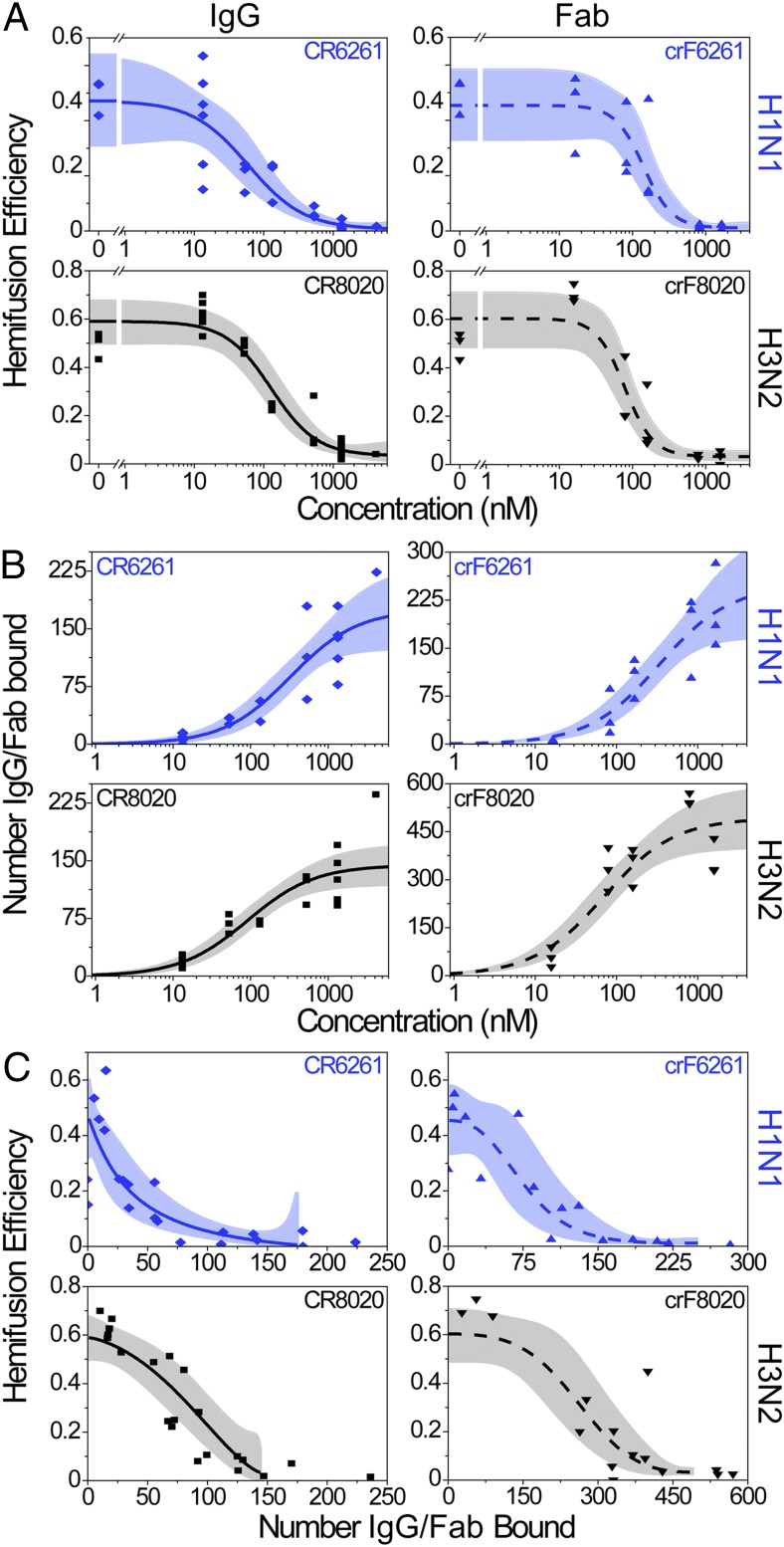
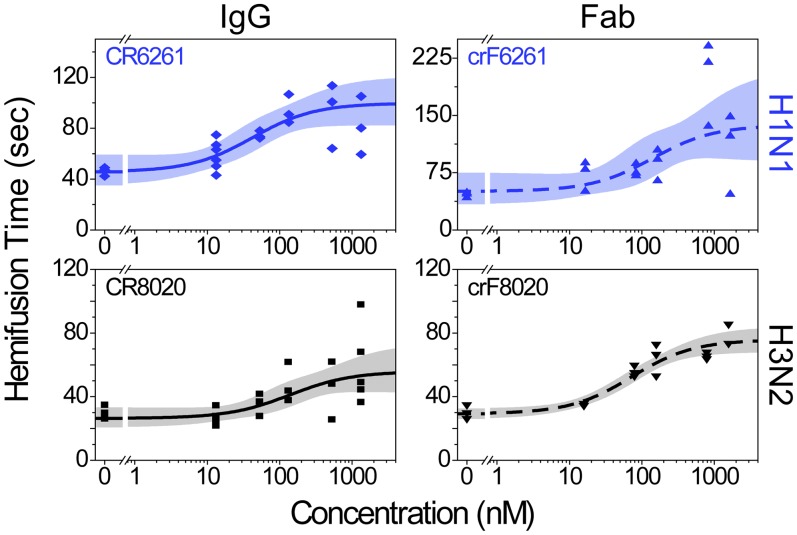
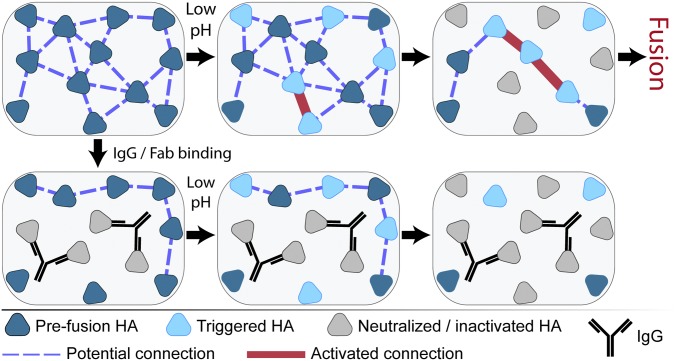
The ability of antibodies binding the influenza hemagglutinin (HA) protein to neutralize viral infectivity is of key importance in the design of next-generation vaccines and for prophylactic and therapeutic use. The two antibodies CR6261 and CR8020 have recently been shown to efficiently neutralize influenza A infection by binding to and inhibiting the influenza A HA protein that is responsible for membrane fusion in the early steps of viral infection. Here, we use single-particle fluorescence microscopy to correlate the number of antibodies or antibody fragments (Fab) bound to an individual virion with the capacity of the same virus particle to undergo membrane fusion. To this end, individual, infectious virus particles bound by fluorescently labeled antibodies/Fab are visualized as they fuse to a planar, supported lipid bilayer. The fluorescence intensity arising from the virus-bound antibodies/Fab is used to determine the number of molecules attached to viral HA while a fluorescent marker in the viral membrane is used to simultaneously obtain kinetic information on the fusion process. We experimentally determine that the stoichiometry required for fusion inhibition by both antibody and Fab leaves large numbers of unbound HA epitopes on the viral surface. Kinetic measurements of the fusion process reveal that those few particles capable of fusion at high antibody/Fab coverage display significantly slower hemifusion kinetics. Overall, our results support a membrane fusion mechanism requiring the stochastic, coordinated action of multiple HA trimers and a model of fusion inhibition by stem-binding antibodies through disruption of this coordinated action.
Keywords: hemagglutinin; influenza; membrane fusion; neutralization stoichiometry; neutralizing antibody.
Conflict of interest statement
Conflict of interest statement: B.B., M.H.K., J.J., C.T., T.K., J.G., R.V., and R.H.E.F. are current employees of the Crucell Vaccine Institute.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
