

Plentiful PtdIns5P from scanty PtdIns(3,5)P2 or from ample PtdIns? PIKfyve-dependent models: Evidence and speculation (response to: DOI 10.1002/bies.201300012)
- PMID: 25404370
- PMCID: PMC4636131
- DOI: 10.1002/bies.201400129
Plentiful PtdIns5P from scanty PtdIns(3,5)P2 or from ample PtdIns? PIKfyve-dependent models: Evidence and speculation (response to: DOI 10.1002/bies.201300012)
Abstract
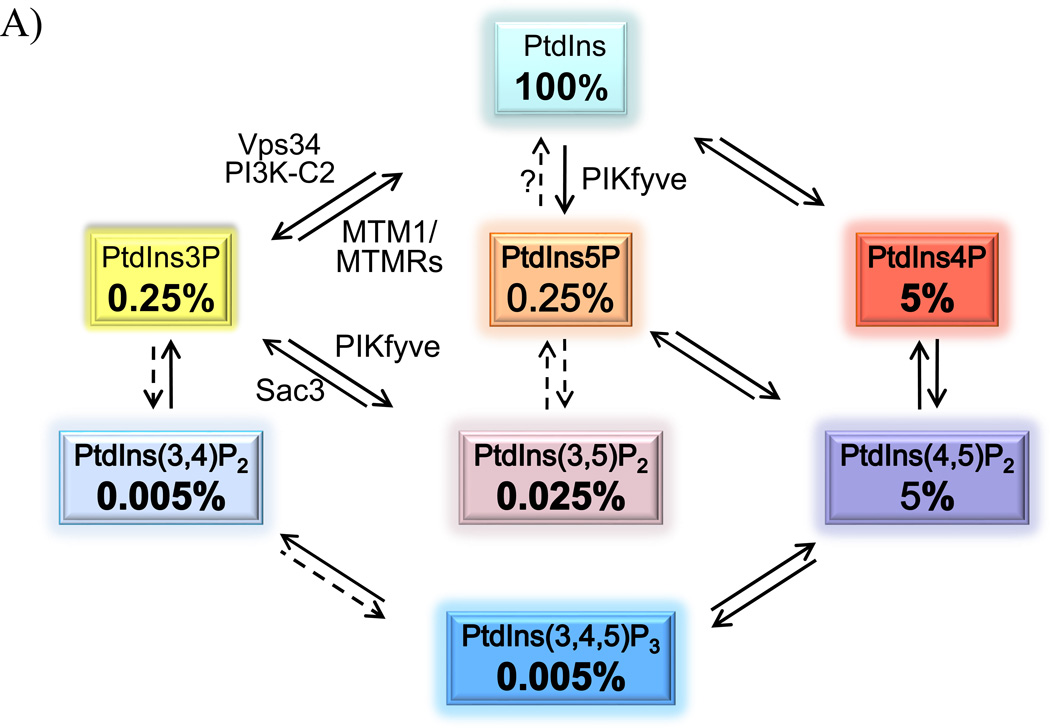
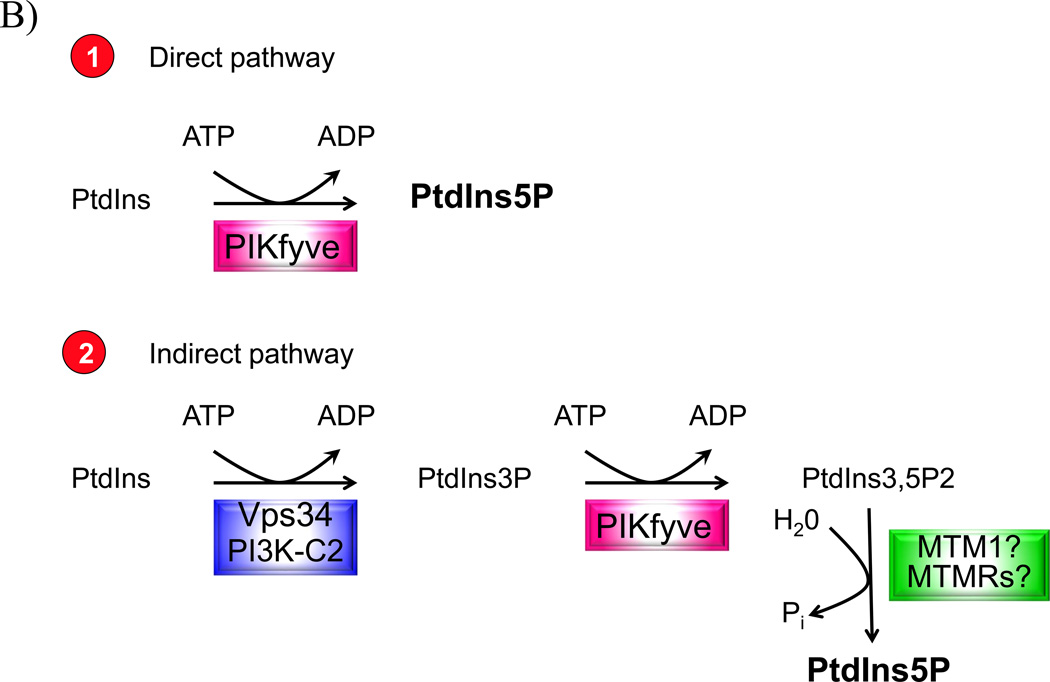
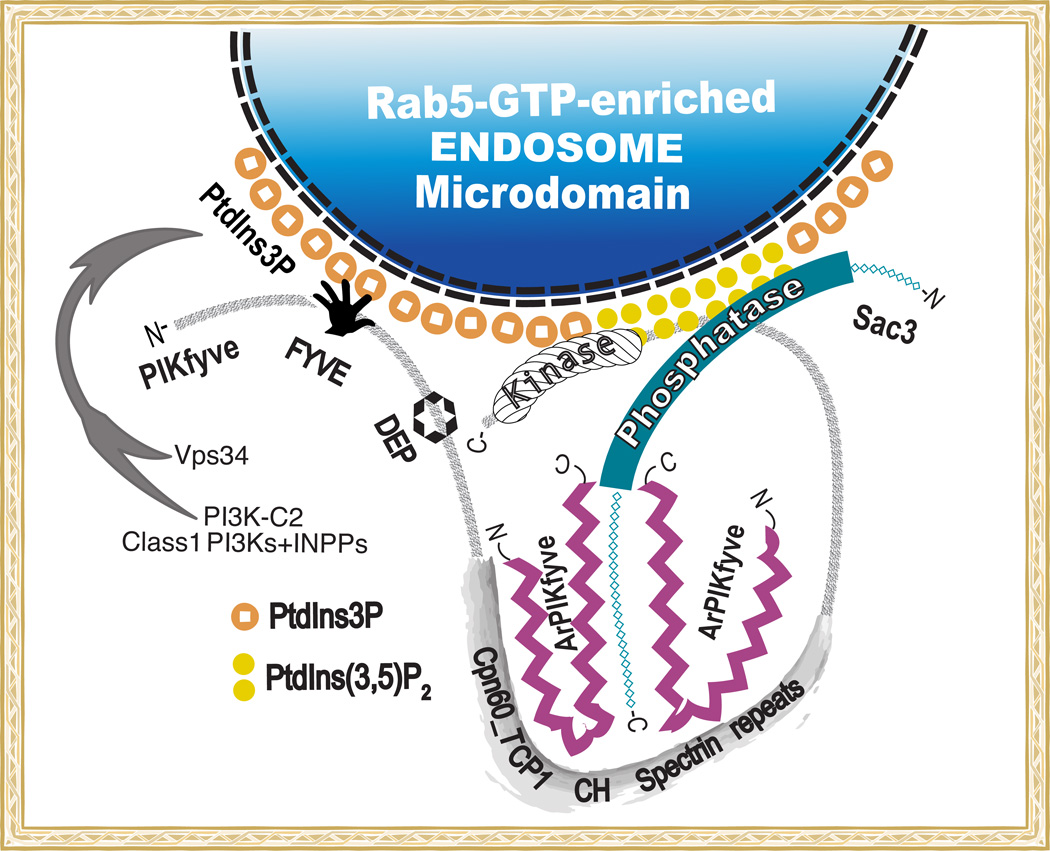
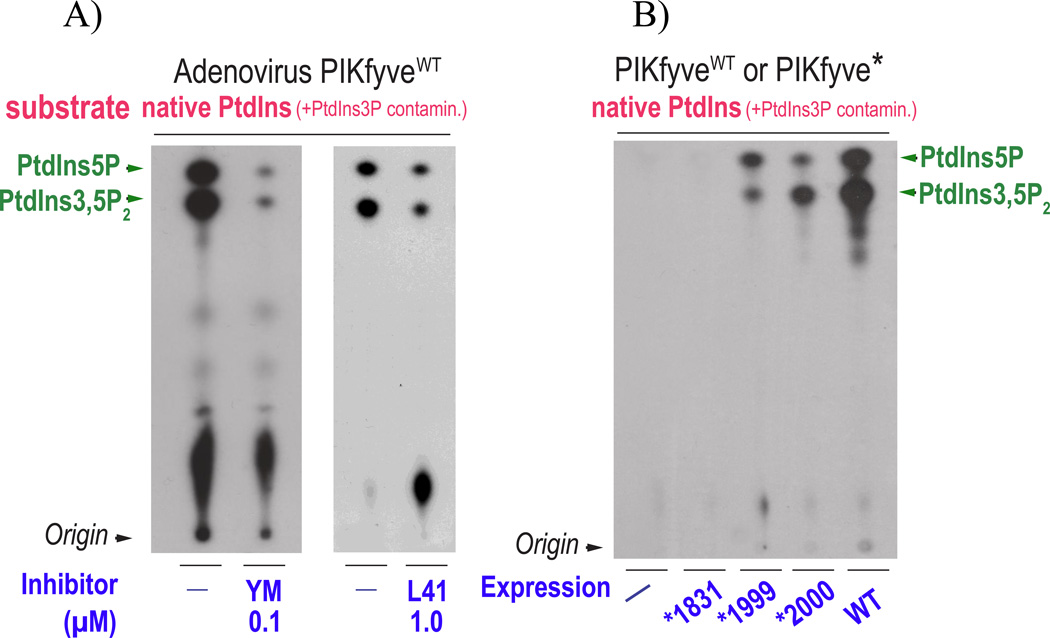
Recently, we have presented data supporting the notion that PIKfyve not only produces the majority of constitutive phosphatidylinositol 5-phosphate (PtdIns5P) in mammalian cells but that it does so through direct synthesis from PtdIns. Another group, albeit obtaining similar data, suggests an alternative pathway whereby the low-abundance PtdIns(3,5)P2 undergoes hydrolysis by unidentified 3-phosphatases, thereby serving as a precursor for most of PtdIns5P. Here, we review the experimental evidence supporting constitutive synthesis of PtdIns5P from PtdIns by PIKfyve. We further emphasize that the experiments presented in support of the alternative pathway are also compatible with a direct mechanism for PIKfyve-catalyzed synthesis of PtdIns5P. While agreeing with the authors that constitutive PtdIns5P could theoretically be produced from PtdIns(3,5)P2 by 3-dephosphorylation, we argue that until direct evidence for such an alternative pathway is obtained, we should adhere to the existing experimental evidence and quantitative considerations, which favor direct PIKfyve-catalyzed synthesis for most constitutive PtdIns5P.
Keywords: 3-phosphatases; PIKfyve; PtdIns(3,5)P2; PtdIns(3,5)P2-PtdIns3P conversion; PtdIns3P; direct PtdIns5P synthesis; phosphoinositides.
© 2015 WILEY Periodicals, Inc.
Conflict of interest statement
Figures
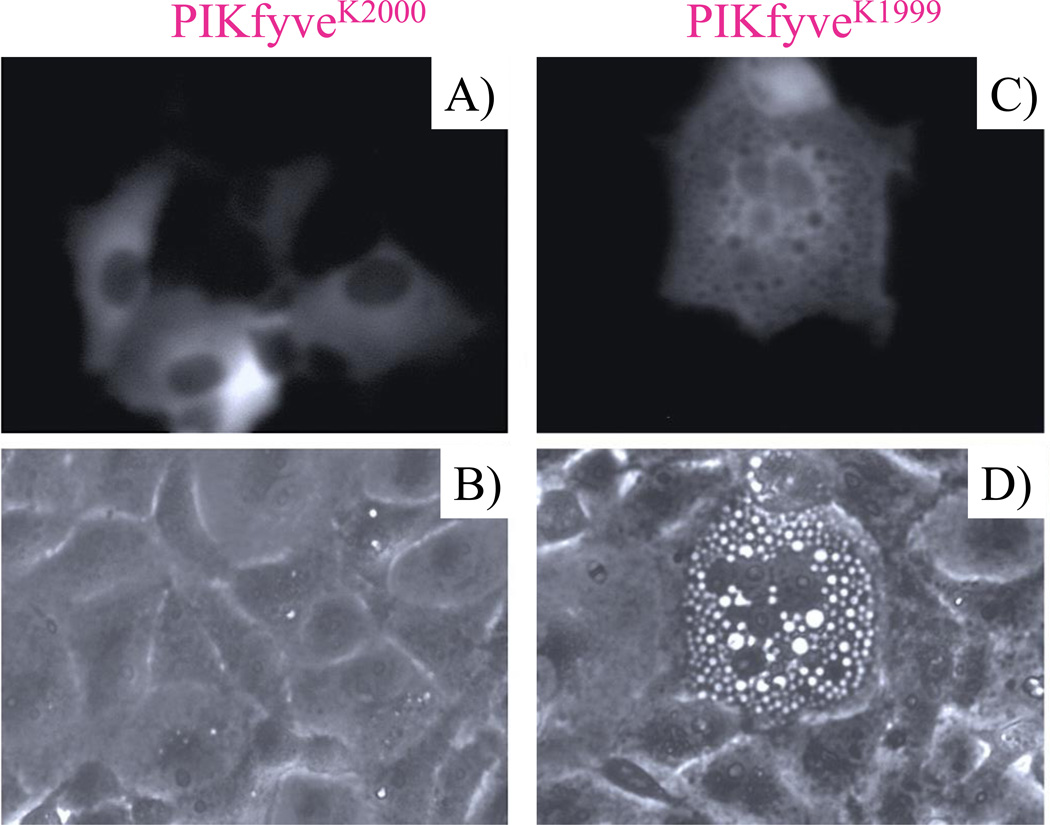
Comment on
-
Phosphatidylinositol 3,5-bisphosphate: low abundance, high significance.Bioessays. 2014 Jan;36(1):52-64. doi: 10.1002/bies.201300012. Epub 2013 Oct 28. Bioessays. 2014. PMID: 24323921 Free PMC article. Review.
References
-
- Di Paolo G, De Camilli P. Phosphoinositides in cell regulation and membrane dynamics. Nature. 2006;443:651–657. - PubMed
-
- Shisheva A. PIKfyve and its lipid products in health and in sickness. Curr Top Microbiol Immunol. 2012;362:127–162. - PubMed
-
- Hakim S, Bertucci MC, Conduit SE, Vuong DL, et al. Inositol polyphosphate phosphatases in human disease. Curr Top Microbiol Immunol. 2012;362:247–314. - PubMed
-
- Viaud J, Lagarrigue F, Ramel D, Allart S, et al. Phosphatidylinositol 5-phosphate regulates invasion through binding and activation of Tiam1. Nat Commun. 2014;5:4080. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
