

Escherichia coli SecG is required for residual export mediated by mutant signal sequences and for SecY-SecE complex stability
- PMID: 25404704
- PMCID: PMC4285974
- DOI: 10.1128/JB.02136-14
Escherichia coli SecG is required for residual export mediated by mutant signal sequences and for SecY-SecE complex stability
Abstract
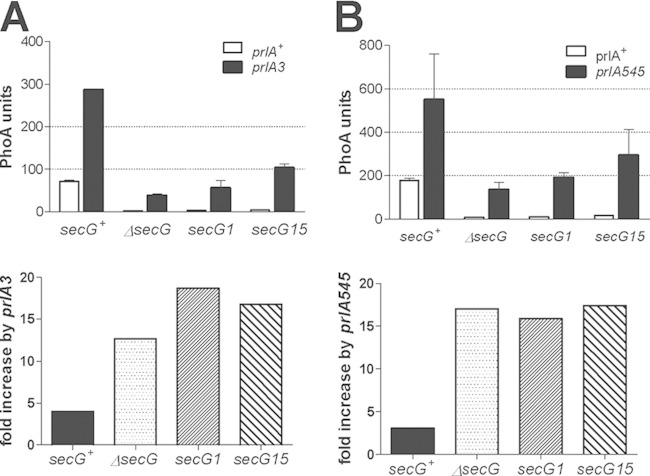
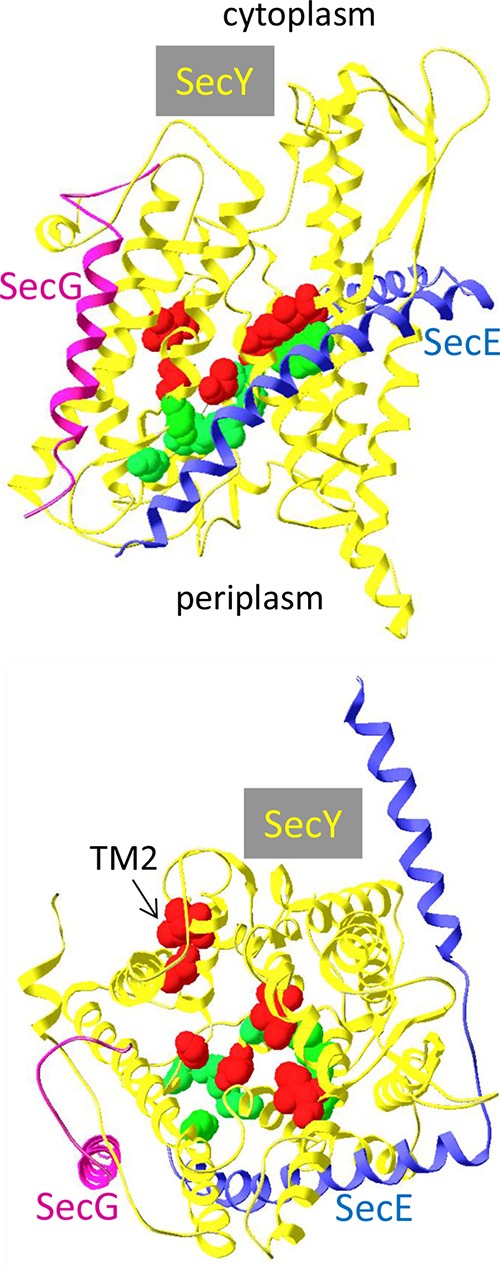
Protein export to the bacterial periplasm is achieved by SecYEG, an inner membrane heterotrimer. SecY and SecE are encoded by essential genes, while SecG is not essential for growth under standard laboratory conditions. Using a quantitative and sensitive export assay, we show that SecG plays a critical role for the residual export mediated by mutant signal sequences; the magnitude of this effect is not proportional to the strength of the export defect. In contrast, export mediated by wild-type signal sequences is only barely retarded in the absence of SecG. When probed with mutant signal sequences, secG loss of function mutations display a phenotype opposite to that of prlA mutations in secY. The analysis of secG and prlA single and double mutant strains shows that the increased export conferred by several prlA alleles is enhanced in the absence of SecG. Several combinations of prlA alleles with a secG deletion cannot be easily constructed. This synthetic phenotype is conditional, indicating that cells can adapt to the presence of both alleles. The biochemical basis of this phenomenon is linked to the stability of the SecYE dimer in solubilized membranes. With prlA alleles that can be normally introduced in a secG deletion strain, SecG has only a limited effect on the stability of the SecYE dimer. With the other prlA alleles, the SecYE dimer can often be detected only in the presence of SecG. A possible role for the maintenance of SecG during evolution is proposed.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures






Similar articles
-
SecG is an auxiliary component of the protein export apparatus of Escherichia coli.Mol Gen Genet. 2000 Feb;263(1):131-6. doi: 10.1007/s004380050039. Mol Gen Genet. 2000. PMID: 10732681
-
Modeling the effects of prl mutations on the Escherichia coli SecY complex.J Bacteriol. 2005 Sep;187(18):6454-65. doi: 10.1128/JB.187.18.6454-6465.2005. J Bacteriol. 2005. PMID: 16159779 Free PMC article.
-
prl mutations in the Escherichia coli secG gene.J Biol Chem. 1997 Feb 14;272(7):4087-93. doi: 10.1074/jbc.272.7.4087. J Biol Chem. 1997. PMID: 9020118
-
Structure, function, and biogenesis of SecY, an integral membrane protein involved in protein export.J Bioenerg Biomembr. 1990 Jun;22(3):353-67. doi: 10.1007/BF00763172. J Bioenerg Biomembr. 1990. PMID: 2202723 Review.
-
The sec and prl genes of Escherichia coli.J Bioenerg Biomembr. 1990 Jun;22(3):291-310. doi: 10.1007/BF00763169. J Bioenerg Biomembr. 1990. PMID: 2202721 Review.
Cited by
-
SecA-Mediated Protein Translocation through the SecYEG Channel.Microbiol Spectr. 2019 Jul;7(4):10.1128/microbiolspec.psib-0028-2019. doi: 10.1128/microbiolspec.PSIB-0028-2019. Microbiol Spectr. 2019. PMID: 31373268 Free PMC article. Review.
-
The way is the goal: how SecA transports proteins across the cytoplasmic membrane in bacteria.FEMS Microbiol Lett. 2018 Jun 1;365(11):fny093. doi: 10.1093/femsle/fny093. FEMS Microbiol Lett. 2018. PMID: 29790985 Free PMC article. Review.
-
AraC Functional Suppressors of Mutations in the C-Terminal Domain of the RpoA Subunit of the Escherichia coli RNA Polymerase.Microorganisms. 2024 Sep 23;12(9):1928. doi: 10.3390/microorganisms12091928. Microorganisms. 2024. PMID: 39338602 Free PMC article.
-
Partially inserted nascent chain unzips the lateral gate of the Sec translocon.EMBO Rep. 2019 Oct 4;20(10):e48191. doi: 10.15252/embr.201948191. Epub 2019 Aug 5. EMBO Rep. 2019. PMID: 31379073 Free PMC article.
-
Fates of Sec, Tat, and YidC Translocases in Mitochondria and Other Eukaryotic Compartments.Mol Biol Evol. 2021 Dec 9;38(12):5241-5254. doi: 10.1093/molbev/msab253. Mol Biol Evol. 2021. PMID: 34436602 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
