

doi: 10.1371/journal.pbio.1001996.
eCollection 2014 Nov.
Mechano-transduction: from molecules to tissues
Affiliations
- PMID: 25405923
- PMCID: PMC4236045
- DOI: 10.1371/journal.pbio.1001996
Item in Clipboard
Mechano-transduction: from molecules to tissues
PLoS Biol.
.
Abstract
External forces play complex roles in cell organization, fate, and homeostasis. Changes in these forces, or how cells respond to them, can result in abnormal embryonic development and diseases in adults. How cells sense and respond to these mechanical stimuli requires an understanding of the biophysical principles that underlie changes in protein conformation and result in alterations in the organization and function of cells and tissues. Here, we discuss mechano-transduction as it applies to protein conformation, cellular organization, and multi-cell (tissue) function.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
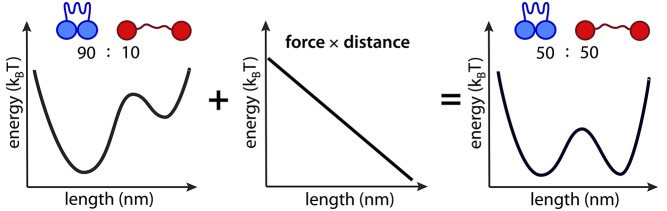
Proteins exist in a conformational equilibrium in which different states are populated according to their relative energies. Mechanical force shifts the equilibrium among pre-existing states. Consider a protein in equilibrium between compact and extended conformations A (blue) and B (red). The corresponding equilibrium constant K = [B]/[A] relates to the free energy difference ΔG between states as: K = exp(-ΔG/kBT), where kB is Boltzmann's constant and T is absolute temperature. An applied load F stabilizes the extended conformation by a mechanical work term of F times distance, δ, leading to a new equilibrium constant K′ = exp[(-ΔG+Fδ/kBT] . In effect, force shifts the energetic balance between the compact and extended states, increasing the amount of protein in the extended conformation by a factor of five in the illustration above. It is useful to remember that since kBT has units of energy, it can be expressed in units of force multiplied by distance and is 4.2 pN nm at physiological temperature. Thus, pN forces acting over nm distances are sufficient to meaningfully shift the equilibrium between conformations.
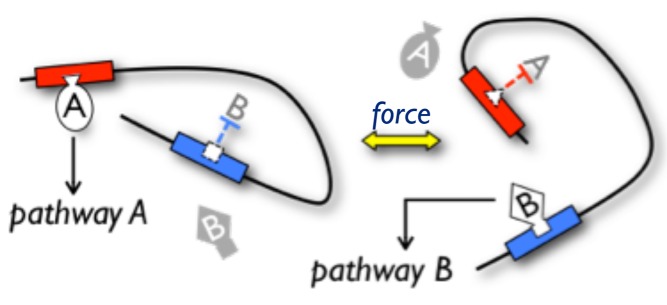
Left: This protein conformation allows protein A to bind and activate its pathway A but blocks binding of protein B, so pathway B is inactive. Right: A change in protein conformation induced by an external force, for example, inhibits the binding of protein A and allows protein B to bind, and so activate, pathway B.

External force is detected by mechano-sensors in the plasma membrane, which link to intracellular adaptors that transmit mechanical signals to targets in the cell. Right: examples of processes and proteins involved. These pathways may exhibit feedback regulation (see main text for details).
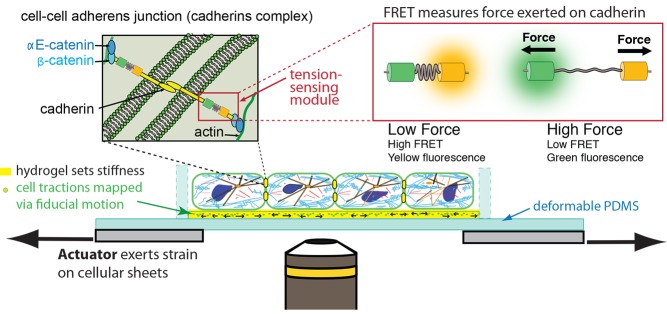
In a monolayer of epithelial cells growing on a hydrogel of known stiffness, cell–cell adherens junctions mechanically link one cell to one another. These junctions comprise transmembrane cadherin proteins that bind to each other in the intracellular space and on the cytoplasmic face of the plasma membrane form ternary complexes with αE- and β-catenin; the molecular identity of the linker to the actin cytoskeleton and other proteins in the tension-sensing module remain unknown. Förster resonance energy transfer (FRET) probes engineered into key proteins allow us to measure quantitatively the molecular-scale forces the cells exert on each other in response to a macroscopic stretch applied by an actuator.
References
-
- Guillot C, Lecuit T (2013) Mechanics of epithelial tissue homeostasis and morphogenesis. Science 340: 1185–1189. - PubMed
-
- Marshall BT, Long M, Piper JW, Yago T, McEver RP, et al. (2003) Direct observation of catch bonds involving cell-adhesion molecules. Nature 423: 190–193. - PubMed
-
- Thomas WE, Vogel V, Sokurenko E (2008) Biophysics of catch bonds. Annu Rev Biophys 37: 399–416. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
