

Two-pore channels provide insight into the evolution of voltage-gated Ca2+ and Na+ channels
- PMID: 25406377
- PMCID: PMC4327855
- DOI: 10.1126/scisignal.2005450
Two-pore channels provide insight into the evolution of voltage-gated Ca2+ and Na+ channels
Abstract
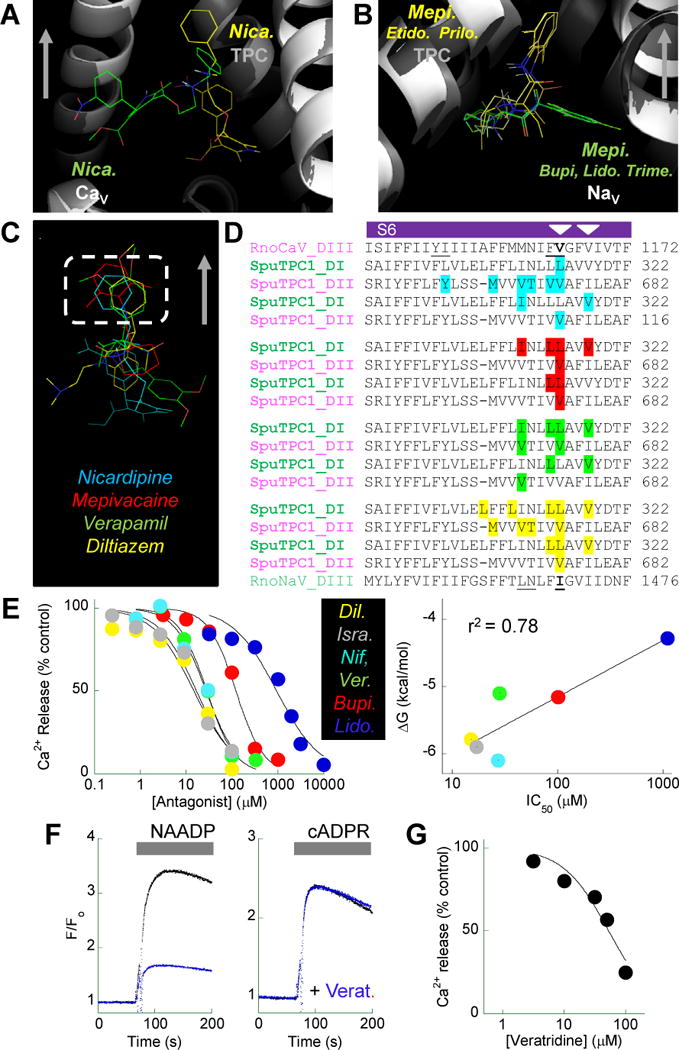
Four-domain voltage-gated Ca(2+) and Na(+) channels (CaV, NaV) underpin nervous system function and likely emerged upon intragenic duplication of a primordial two-domain precursor. To investigate if two-pore channels (TPCs) may represent an intermediate in this evolutionary transition, we performed molecular docking simulations with a homology model of TPC1, which suggested that the pore region could bind antagonists of CaV or NaV. CaV or NaV antagonists blocked NAADP (nicotinic acid adenine dinucleotide phosphate)-evoked Ca(2+) signals in sea urchin egg preparations and in intact cells that overexpressed TPC1. By sequence analysis and inspection of the model, we predicted a noncanonical selectivity filter in animal TPCs in which the carbonyl groups of conserved asparagine residues are positioned to coordinate cations. In contrast, a distinct clade of TPCs [TPCR (for TPC-related)] in several unicellular species had ion selectivity filters with acidic residues more akin to CaV. TPCRs were predicted to interact strongly with CaV antagonists. Our data suggest that acquisition of a "blueprint" pharmacological profile and changes in ion selectivity within four-domain voltage-gated ion channels may have predated intragenic duplication of an ancient two-domain ancestor.
Copyright © 2014, American Association for the Advancement of Science.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
