

A pneumocyte-macrophage paracrine lipid axis drives the lung toward fibrosis
- PMID: 25409201
- PMCID: PMC4566113
- DOI: 10.1165/rcmb.2014-0343OC
A pneumocyte-macrophage paracrine lipid axis drives the lung toward fibrosis
Abstract
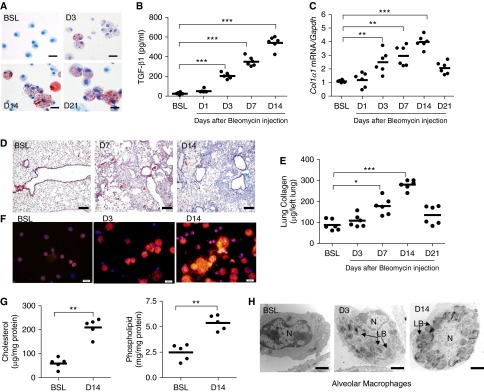
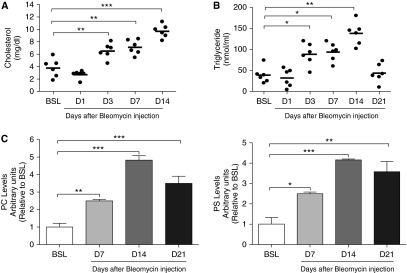
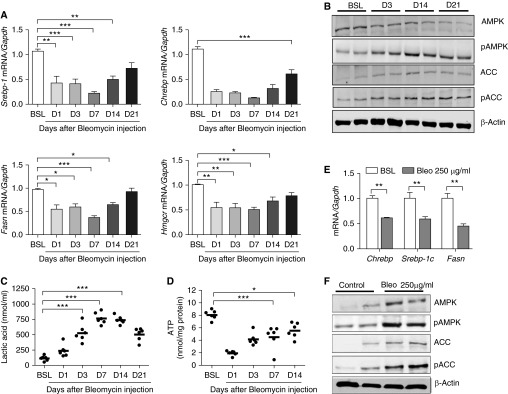
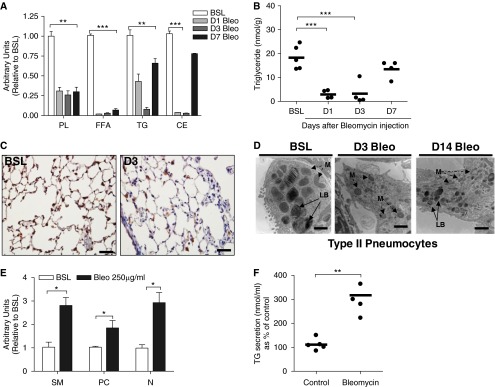
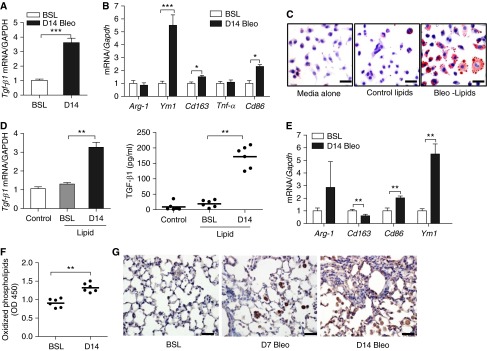
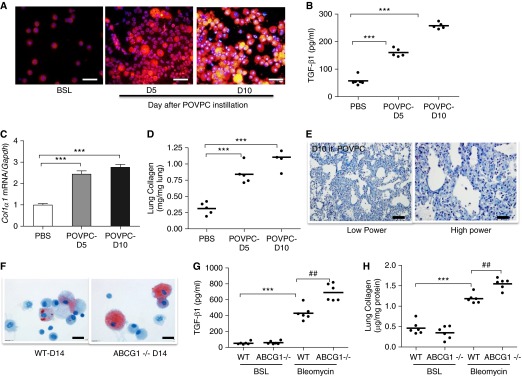
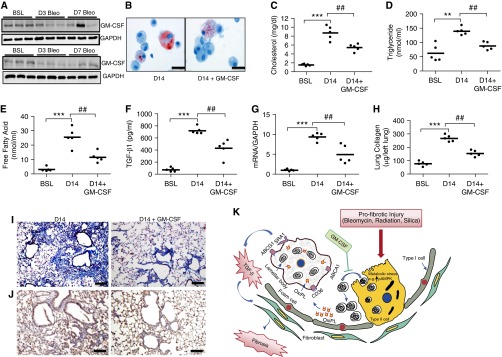
Lipid-laden macrophages, or "foam cells," are observed in the lungs of patients with fibrotic lung disease, but their contribution to disease pathogenesis remains unexplored. Here, we demonstrate that fibrosis induced by bleomycin, silica dust, or thoracic radiation promotes early and sustained accumulation of foam cells in the lung. In the bleomycin model, we show that foam cells arise from neighboring alveolar epithelial type II cells, which respond to injury by dumping lipids into the distal airspaces of the lungs. We demonstrate that oxidized phospholipids accumulate within alveolar macrophages (AMs) after bleomycin injury and that murine and human AMs treated with oxidized phosphatidylcholine (oxPc) become polarized along an M2 phenotype and display enhanced production of transforming growth factor-β1. The direct instillation of oxPc into the mouse lung induces foam cell formation and triggers a severe fibrotic reaction. Further, we show that reducing pulmonary lipid clearance by targeted deletion of the lipid efflux transporter ATP-binding cassette subfamily G member 1 increases foam cell formation and worsens lung fibrosis after bleomycin. Conversely, we found that treatment with granulocyte-macrophage colony-stimulating factor attenuates fibrotic responses, at least in part through its ability to decrease AM lipid accumulation. In summary, this work describes a novel mechanism leading to foam cell formation in the mouse lung and suggests that strategies aimed at blocking foam cell formation might be effective for treating fibrotic lung disorders.
Keywords: alveolar macrophages; foam cells; oxidized phospholipids; pulmonary fibrosis; type II pneumocytes.
Figures
References
-
- van Moorsel CH, van Oosterhout MF, Barlo NP, de Jong PA, van der Vis JJ, Ruven HJ, van Es HW, van den Bosch JM, Grutters JC. Surfactant protein C mutations are the basis of a significant portion of adult familial pulmonary fibrosis in a dutch cohort. Am J Respir Crit Care Med. 2010;182:1419–1425. - PubMed
-
- Walters DM, Cho HY, Kleeberger SR. Oxidative stress and antioxidants in the pathogenesis of pulmonary fibrosis: a potential role for Nrf2. Antioxid Redox Signal. 2008;10:321–332. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
