

Differentially methylated plasticity genes in the amygdala of young primates are linked to anxious temperament, an at risk phenotype for anxiety and depressive disorders
- PMID: 25411484
- PMCID: PMC4236392
- DOI: 10.1523/JNEUROSCI.3338-14.2014
Differentially methylated plasticity genes in the amygdala of young primates are linked to anxious temperament, an at risk phenotype for anxiety and depressive disorders
Abstract
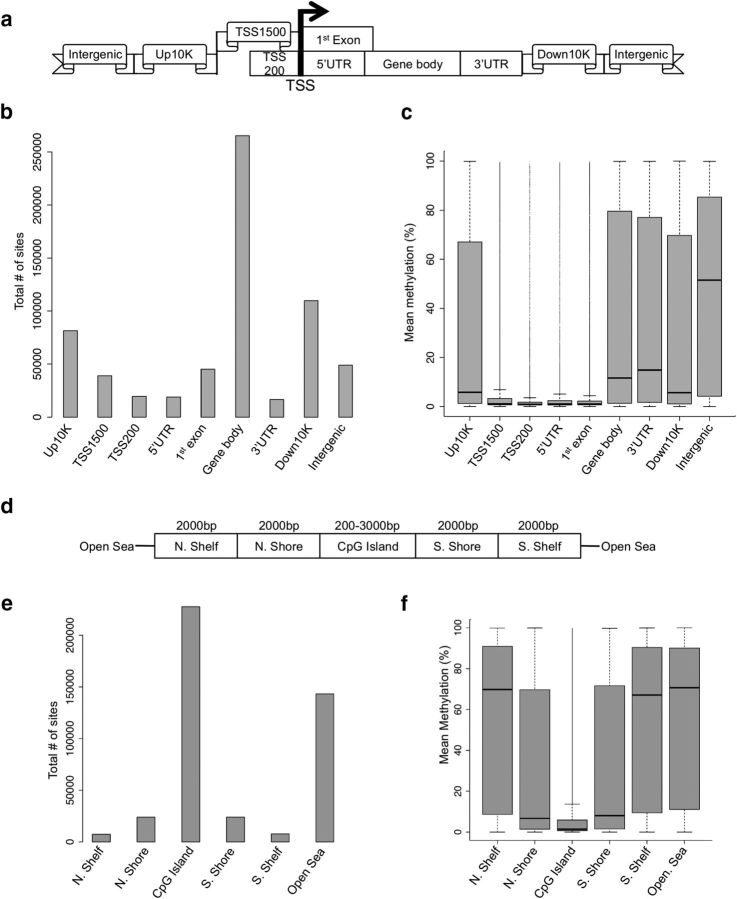
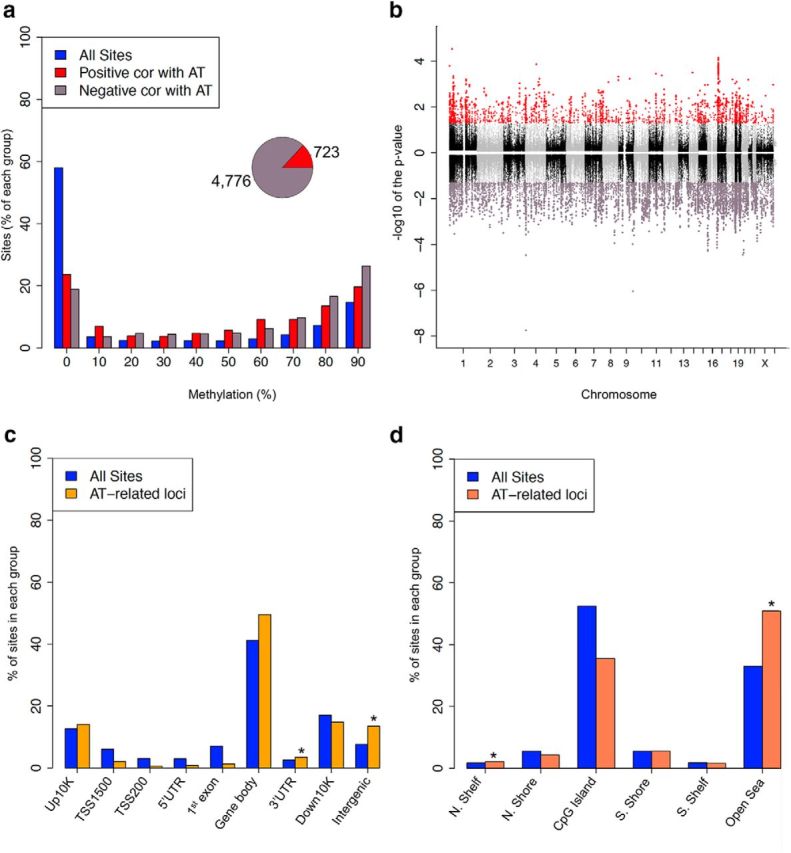
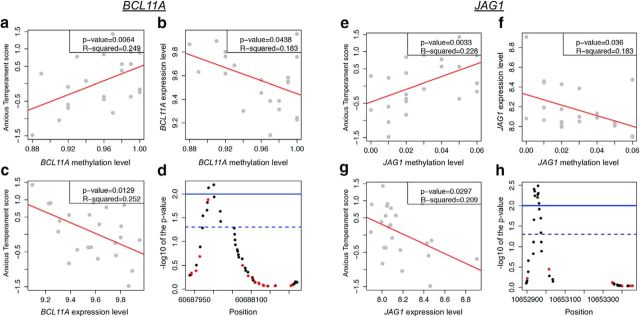
Children with an anxious temperament (AT) are at a substantially increased risk to develop anxiety and depression. The young rhesus monkey is ideal for studying the origin of human AT because it shares with humans the genetic, neural, and phenotypic underpinnings of complex social and emotional functioning. Heritability, functional imaging, and gene expression studies of AT in young monkeys revealed that the central nucleus of the amygdala (Ce) is a key environmentally sensitive substrate of this at risk phenotype. Because epigenetic marks (e.g., DNA methylation) can be modulated by environmental stimuli, these data led us to hypothesize a role for DNA methylation in the development of AT. To test this hypothesis, we used reduced representation bisulfite sequencing to examine the cross-sectional genome-wide methylation levels in the Ce of 23 age-matched monkeys (1.3 ± 0.2 years) phenotyped for AT. Because AT reflects a continuous trait-like variable, we used an analytical approach that is consistent with this biology to identify genes in the Ce with methylation patterns that predict AT. Expression data from the Ce of these same monkeys were then used to find differentially methylated candidates linked to altered gene regulation. Two genes particularly relevant to the AT phenotype were BCL11A and JAG1. These transcripts have well-defined roles in neurodevelopmental processes, including neurite arborization and the regulation of neurogenesis. Together, these findings represent a critical step toward understanding the effects of early environment on the neuromolecular mechanisms that underlie the risk to develop anxiety and depressive disorders.
Keywords: BCL11A; Ce; DNA methylation; JAG1; anxious temperament.
Copyright © 2014 the authors 0270-6474/14/3415548-09$15.00/0.
Figures
References
-
- Davidson RJ, Fox AS, Kalin NH. Neural bases of emotion regulation in non-human primates and humans. New York: Guildford; 2006.
-
- Dempster EL, Pidsley R, Schalkwyk LC, Owens S, Georgiades A, Kane F, Kalidindi S, Picchioni M, Kravariti E, Toulopoulou T, Murray RM, Mill J. Disease-associated epigenetic changes in monozygotic twins discordant for schizophrenia and bipolar disorder. Hum Mol Genet. 2011;20:4786–4796. doi: 10.1093/hmg/ddr416. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases