

SAS-1 is a C2 domain protein critical for centriole integrity in C. elegans
- PMID: 25412110
- PMCID: PMC4238951
- DOI: 10.1371/journal.pgen.1004777
SAS-1 is a C2 domain protein critical for centriole integrity in C. elegans
Abstract
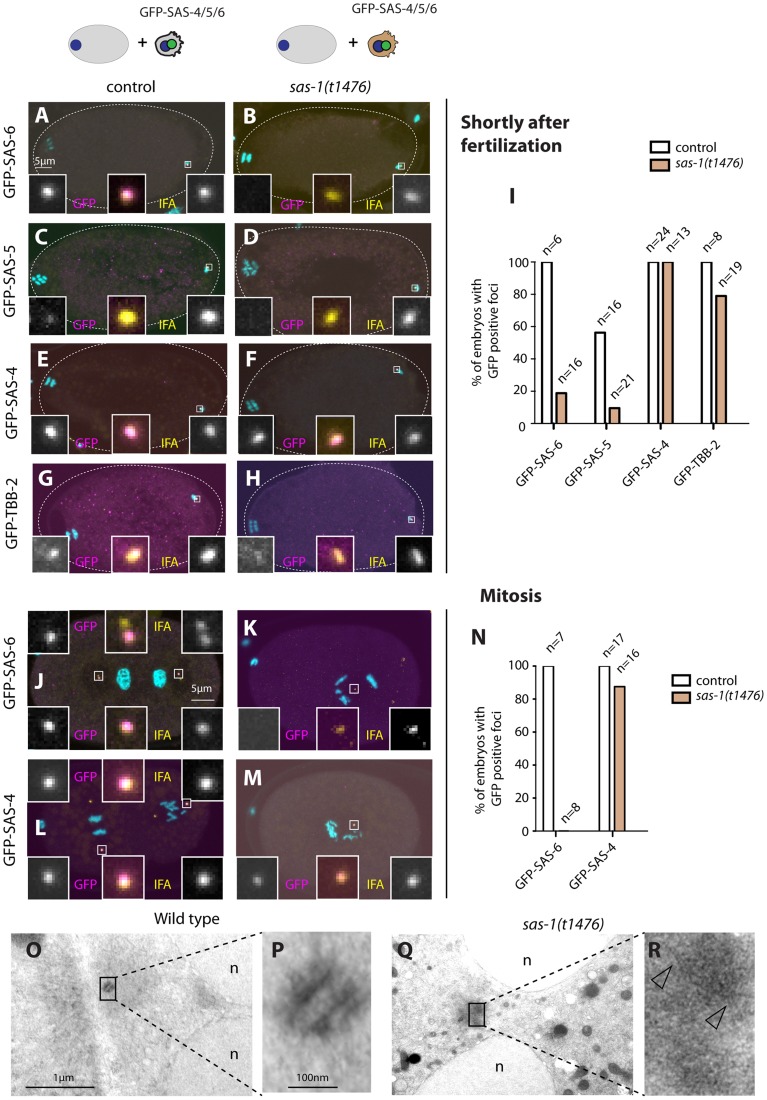
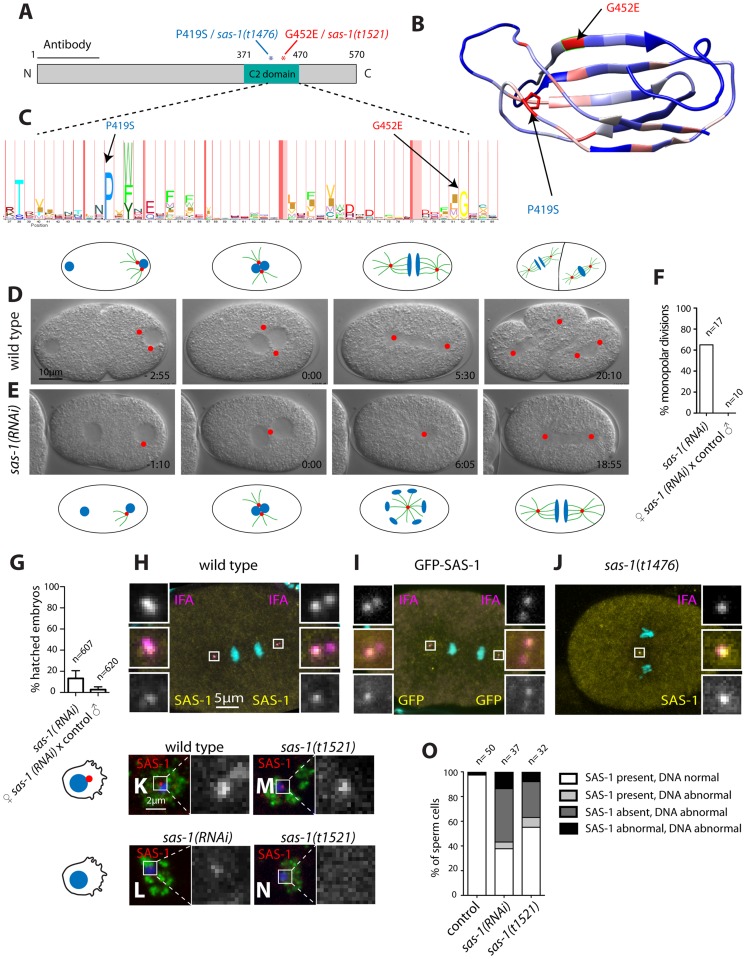
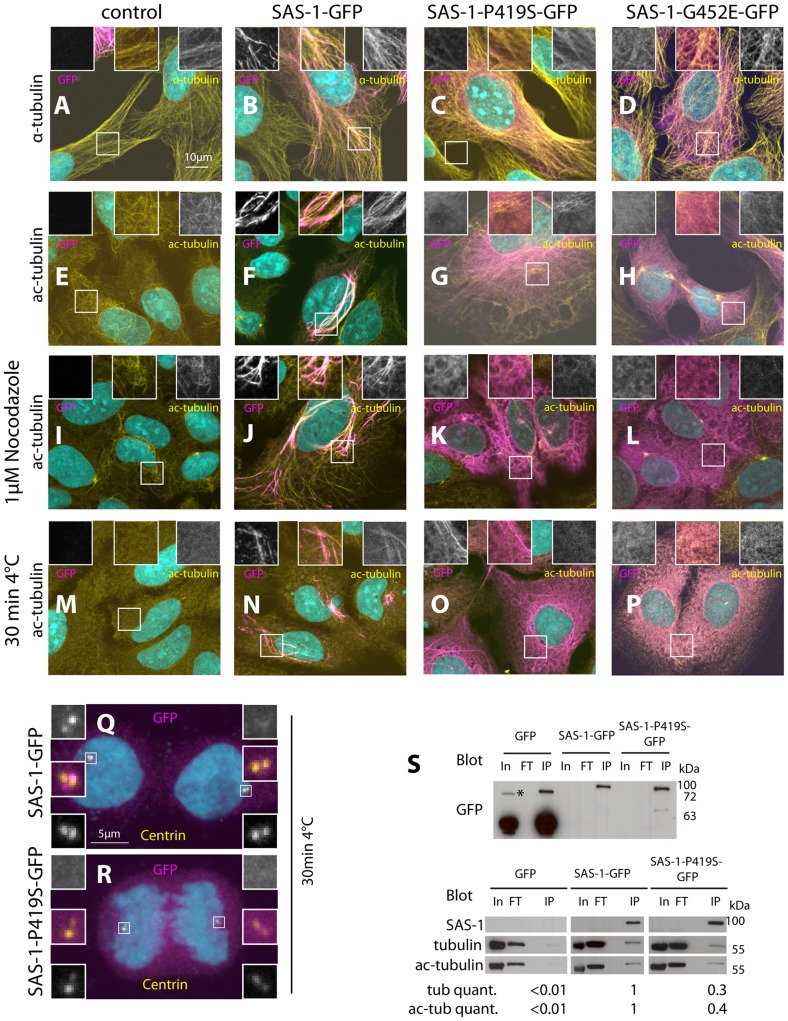
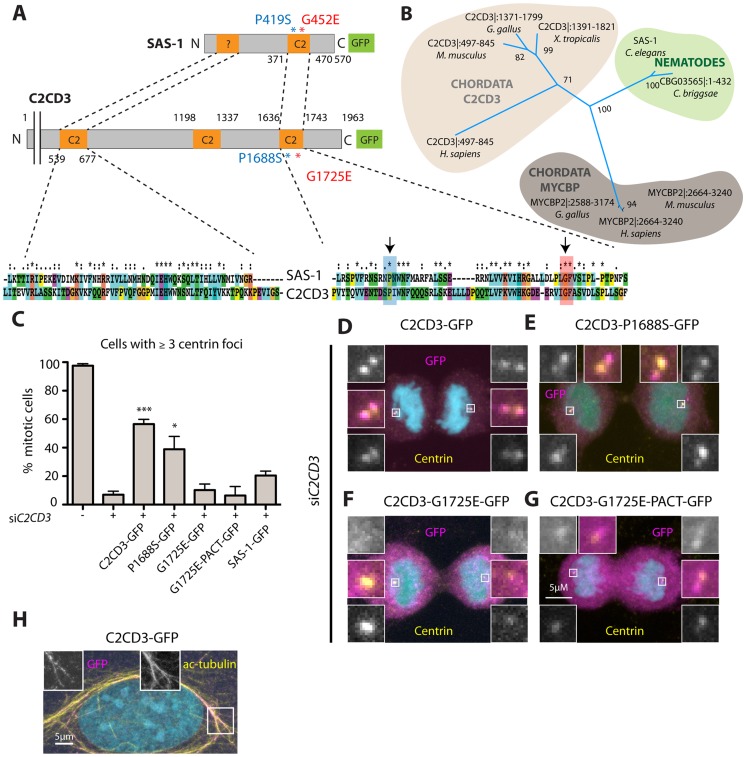
Centrioles are microtubule-based organelles important for the formation of cilia, flagella and centrosomes. Despite progress in understanding the underlying assembly mechanisms, how centriole integrity is ensured is incompletely understood, including in sperm cells, where such integrity is particularly critical. We identified C. elegans sas-1 in a genetic screen as a locus required for bipolar spindle assembly in the early embryo. Our analysis reveals that sperm-derived sas-1 mutant centrioles lose their integrity shortly after fertilization, and that a related defect occurs when maternal sas-1 function is lacking. We establish that sas-1 encodes a C2 domain containing protein that localizes to centrioles in C. elegans, and which can bind and stabilize microtubules when expressed in human cells. Moreover, we uncover that SAS-1 is related to C2CD3, a protein required for complete centriole formation in human cells and affected in a type of oral-facial-digital (OFD) syndrome.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Interaction between the Caenorhabditis elegans centriolar protein SAS-5 and microtubules facilitates organelle assembly.Mol Biol Cell. 2018 Mar 15;29(6):722-735. doi: 10.1091/mbc.E17-06-0412. Epub 2018 Jan 24. Mol Biol Cell. 2018. PMID: 29367435 Free PMC article.
-
Centriolar SAS-5 is required for centrosome duplication in C. elegans.Nat Cell Biol. 2004 Jul;6(7):656-64. doi: 10.1038/ncb1146. Nat Cell Biol. 2004. PMID: 15232593
-
Sequential protein recruitment in C. elegans centriole formation.Curr Biol. 2006 Sep 19;16(18):1844-9. doi: 10.1016/j.cub.2006.07.059. Curr Biol. 2006. PMID: 16979563
-
From zero to many: control of centriole number in development and disease.Traffic. 2009 May;10(5):482-98. doi: 10.1111/j.1600-0854.2009.00905.x. Traffic. 2009. PMID: 19416494 Review.
-
The Rise of the Cartwheel: Seeding the Centriole Organelle.Bioessays. 2018 Apr;40(4):e1700241. doi: 10.1002/bies.201700241. Epub 2018 Mar 6. Bioessays. 2018. PMID: 29508910 Review.
Cited by
-
Centriole elimination during Caenorhabditis elegans oogenesis initiates with loss of the central tube protein SAS-1.EMBO J. 2023 Dec 11;42(24):e115076. doi: 10.15252/embj.2023115076. Epub 2023 Nov 21. EMBO J. 2023. PMID: 37987153 Free PMC article.
-
Towards understanding centriole elimination.Open Biol. 2023 Nov;13(11):230222. doi: 10.1098/rsob.230222. Epub 2023 Nov 15. Open Biol. 2023. PMID: 37963546 Free PMC article. Review.
-
Centriolar Protein C2cd3 Is Required for Craniofacial Development.Front Cell Dev Biol. 2021 Jun 15;9:647391. doi: 10.3389/fcell.2021.647391. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34211969 Free PMC article.
-
A distal centriolar protein network controls organelle maturation and asymmetry.Nat Commun. 2018 Sep 26;9(1):3938. doi: 10.1038/s41467-018-06286-y. Nat Commun. 2018. PMID: 30258116 Free PMC article.
-
A modified TurboID approach identifies tissue-specific centriolar components in C. elegans.PLoS Genet. 2022 Apr 20;18(4):e1010150. doi: 10.1371/journal.pgen.1010150. eCollection 2022 Apr. PLoS Genet. 2022. PMID: 35442950 Free PMC article.
References
-
- Gönczy P (2012) Towards a molecular architecture of centriole assembly. Nat Rev Mol Cell Biol 13: 425–435. - PubMed
-
- Kemp CA, Kopish KR, Zipperlen P, Ahringer J, O'Connell KF (2004) Centrosome maturation and duplication in C. elegans require the coiled-coil protein SPD-2. Dev Cell 6: 511–523. - PubMed
-
- Pelletier L, Ozlü N, Hannak E, Cowan C, Habermann B, et al. (2004) The Caenorhabditis elegans centrosomal protein SPD-2 is required for both pericentriolar material recruitment and centriole duplication. Curr Biol 14: 863–873. - PubMed
-
- O'Connell KF, Caron C, Kopish KR, Hurd DD, Kemphues KJ, et al. (2001) The C. elegans zyg-1 gene encodes a regulator of centrosome duplication with distinct maternal and paternal roles in the embryo. Cell 105: 547–558. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
