

N-terminal acetylation and replicative age affect proteasome localization and cell fitness during aging
- PMID: 25413350
- PMCID: PMC4282048
- DOI: 10.1242/jcs.157354
N-terminal acetylation and replicative age affect proteasome localization and cell fitness during aging
Abstract
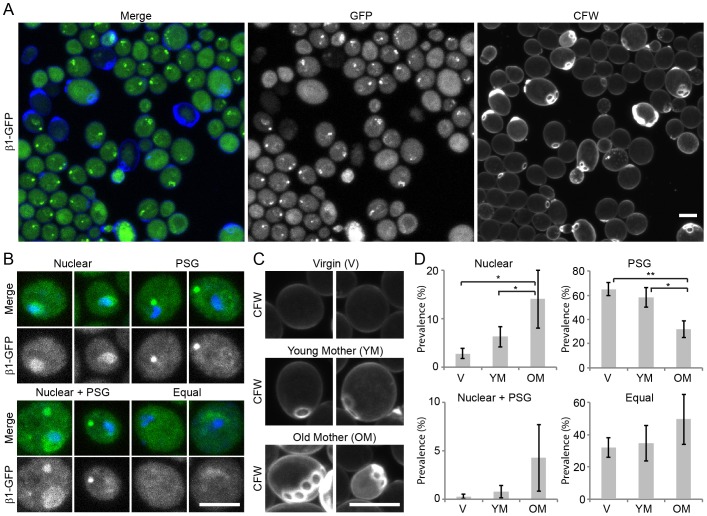
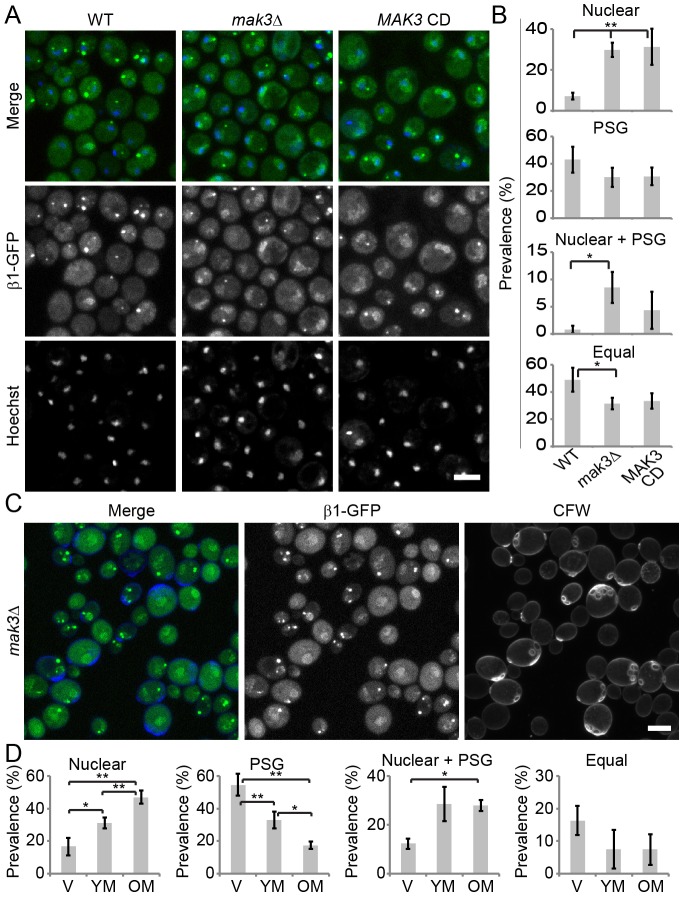
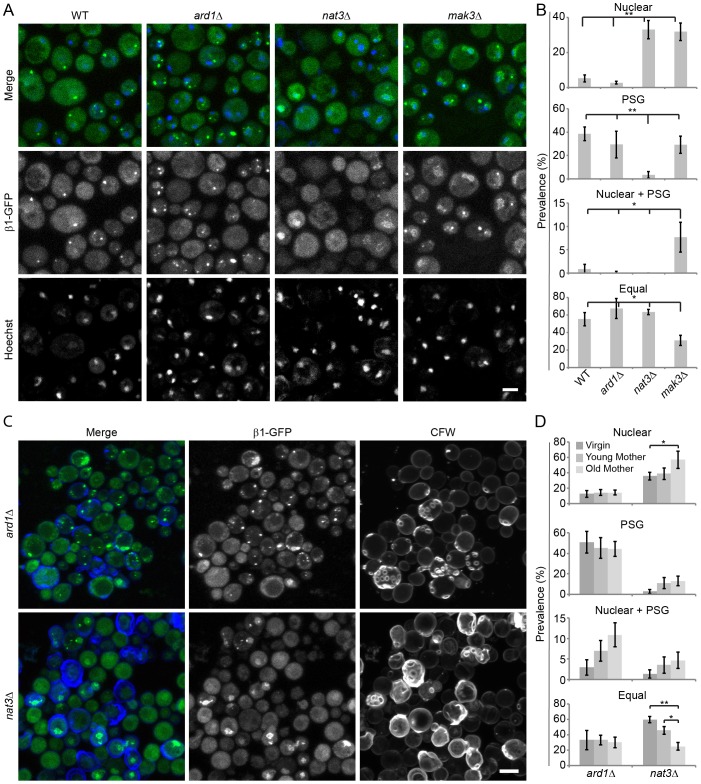
Specific degradation of proteins is essential for virtually all cellular processes and is carried out predominantly by the proteasome. The proteasome is important for clearance of damaged cellular proteins. Damaged proteins accumulate over time and excess damaged proteins can aggregate and induce the death of old cells. In yeast, the localization of the proteasome changes dramatically during aging, possibly in response to altered proteasome activity requirements. We followed two key parameters of this process: the distribution of proteasomes in nuclear and cytosolic compartments, and the formation of cytoplasmic aggregate-like structures called proteasome storage granules (PSGs). Whereas replicative young cells efficiently relocalized proteasomes from the nucleus to the cytoplasm and formed PSGs, replicative old cells were less efficient in relocalizing the proteasome and had less PSGs. By using a microscopy-based genome-wide screen, we identified genetic factors involved in these processes. Both relocalization of the proteasome and PSG formation were affected by two of the three N-acetylation complexes. These N-acetylation complexes also had different effects on the longevity of cells, indicating that each N-acetylation complex has different roles in proteasome location and aging.
Keywords: Aging; Intracellular location; N-acetylation; Proteasome; Replicative age.
© 2015. Published by The Company of Biologists Ltd.
Figures


References
-
- Aragon A. D., Rodriguez A. L., Meirelles O., Roy S., Davidson G. S., Tapia P. H., Allen C., Joe R., Benn D., Werner-Washburne M. (2008). Characterization of differentiated quiescent and nonquiescent cells in yeast stationary-phase cultures. Mol. Biol. Cell 19, 1271–1280 10.1091/mbc.E07-07-0666 - DOI - PMC - PubMed
-
- Arnesen T., Van Damme P., Polevoda B., Helsens K., Evjenth R., Colaert N., Varhaug J. E., Vandekerckhove J., Lillehaug J. R., Sherman F. et al. (2009). Proteomics analyses reveal the evolutionary conservation and divergence of N-terminal acetyltransferases from yeast and humans. Proc. Natl. Acad. Sci. USA 106, 8157–8162 10.1073/pnas.0901931106 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
