

Deciphering the fine details of c1 assembly and activation mechanisms: "mission impossible"?
- PMID: 25414705
- PMCID: PMC4222235
- DOI: 10.3389/fimmu.2014.00565
Deciphering the fine details of c1 assembly and activation mechanisms: "mission impossible"?
Abstract
The classical complement pathway is initiated by the large (~800 kDa) and flexible multimeric C1 complex. Its catalytic function is triggered by the proteases hetero-tetramer C1r2s2, which is associated to the C1q sensing unit, a complex assembly of 18 chains built as a hexamer of heterotrimers. Initial pioneering studies gained insights into the main architectural principles of the C1 complex. A dissection strategy then provided the high-resolution structures of its main functional and/or structural building blocks, as well as structural details on some key protein-protein interactions. These past and current discoveries will be briefly summed up in order to address the question of what is still ill-defined. On a functional point of view, the main molecular determinants of C1 activation and its tight control will be delineated. The current perspective remains to decipher how C1 really works and is controlled in vivo, both in normal and pathological settings.
Keywords: C1 complex; C1q; C1r; C1s; X-ray structures; classical complement pathway; complement-dependent cytotoxicity; serine protease activation.
Figures
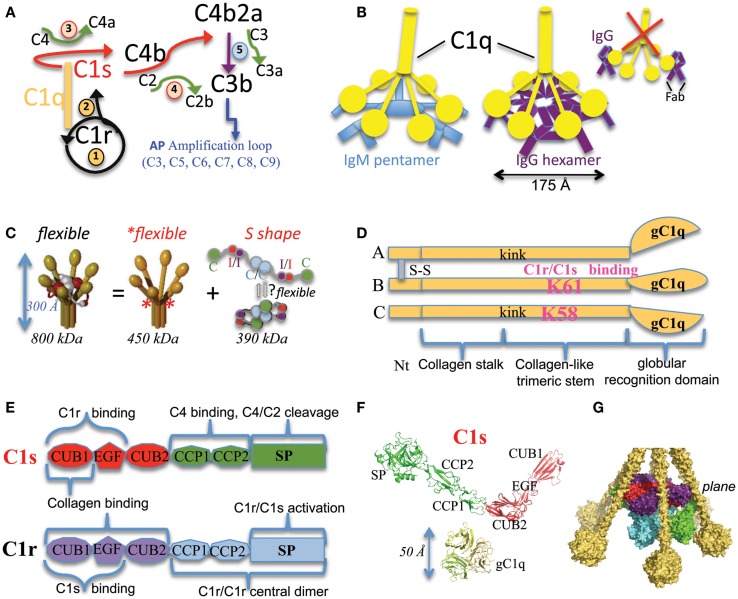
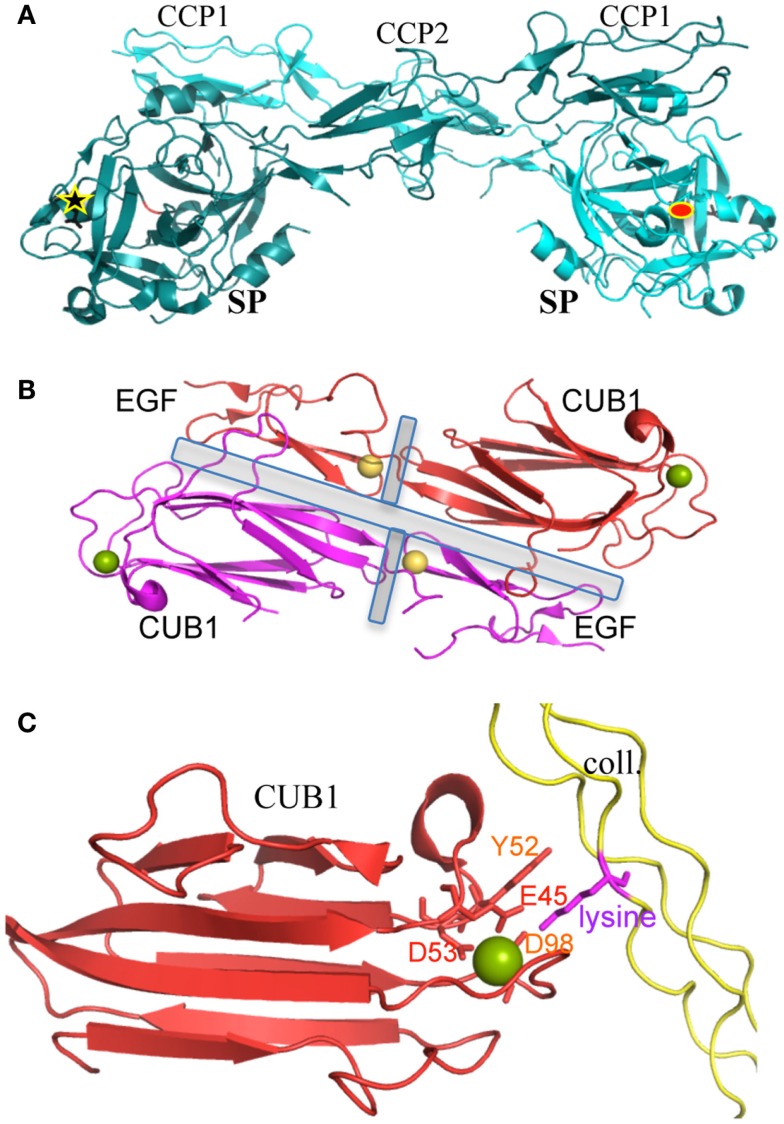
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous