

Integrin αDβ2 (CD11d/CD18) is expressed by human circulating and tissue myeloid leukocytes and mediates inflammatory signaling
- PMID: 25415295
- PMCID: PMC4240710
- DOI: 10.1371/journal.pone.0112770
Integrin αDβ2 (CD11d/CD18) is expressed by human circulating and tissue myeloid leukocytes and mediates inflammatory signaling
Abstract
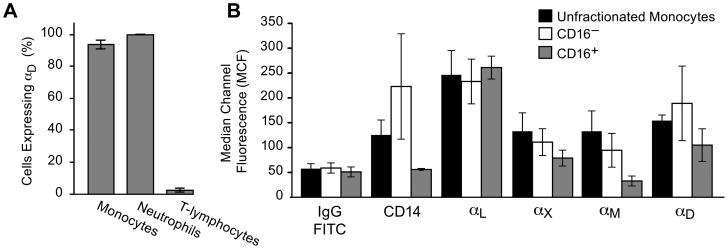
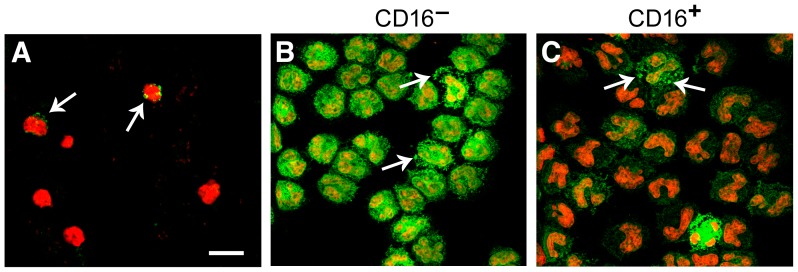
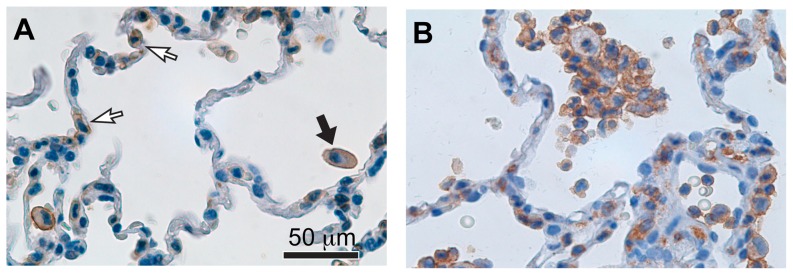
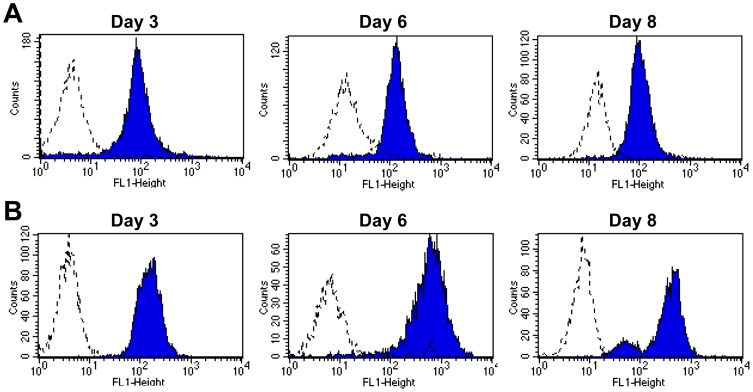
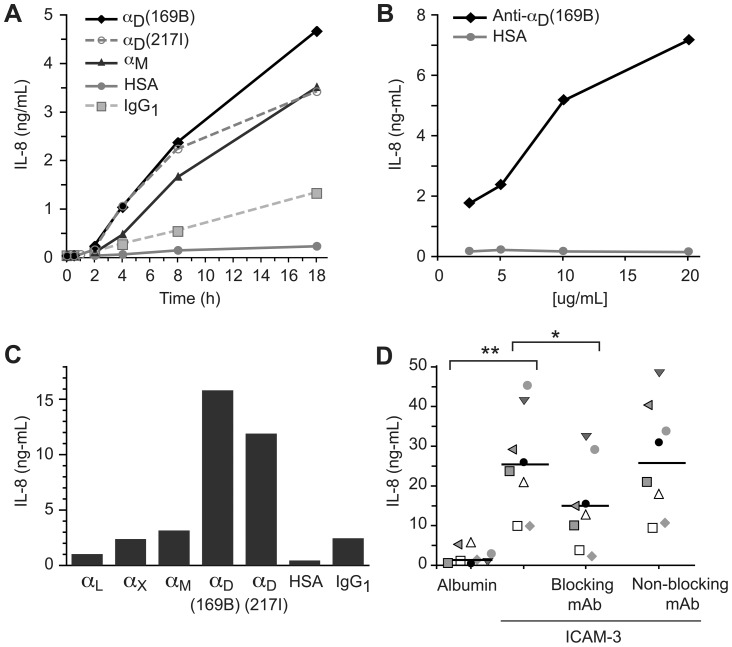
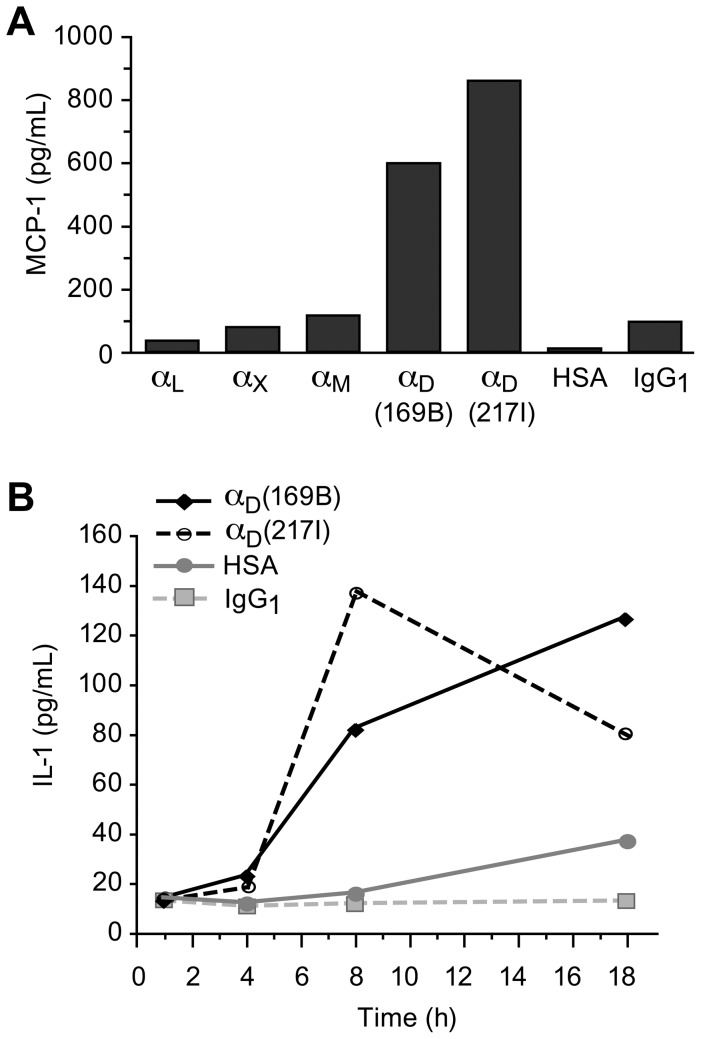
Integrin α(D)β(2) is the most recently identified member of the leukocyte, or β(2), subfamily of integrin heterodimers. Its distribution and functions on human leukocytes have not been clearly defined and are controversial. We examined these issues and found that α(D)β(2) is prominently expressed by leukocytes in whole blood from healthy human subjects, including most polymorphonuclear leukocytes and monocytes. We also found that α(D)β(2) is displayed by leukocytes in the alveoli of uninjured and inflamed human lungs and by human monocyte-derived macrophages and dendritic cells, indicating broad myeloid expression. Using freshly-isolated human monocytes, we found that α(D)β(2) delivers outside-in signals to pathways that regulate cell spreading and gene expression. Screening expression analysis followed by validation of candidate transcripts demonstrated that engagement of α(D)β(2) induces mRNAs encoding inflammatory chemokines and cytokines and secretion of their protein products. Thus, α(D)β(2) is a major member of the integrin repertoire of both circulating and tissue myeloid leukocytes in humans. Its broad expression and capacity for outside-in signaling indicate that it is likely to have important functions in clinical syndromes of infection, inflammation, and tissue injury.
Conflict of interest statement
Figures
Similar articles
-
The Role of Integrins αMβ2 (Mac-1, CD11b/CD18) and αDβ2 (CD11d/CD18) in Macrophage Fusion.Am J Pathol. 2016 Aug;186(8):2105-2116. doi: 10.1016/j.ajpath.2016.04.001. Epub 2016 Jun 14. Am J Pathol. 2016. PMID: 27315778 Free PMC article.
-
Integrin αDβ2 (CD11d/CD18) Modulates Leukocyte Accumulation, Pathogen Clearance, and Pyroptosis in Experimental Salmonella Typhimurium Infection.Front Immunol. 2018 May 24;9:1128. doi: 10.3389/fimmu.2018.01128. eCollection 2018. Front Immunol. 2018. PMID: 29881383 Free PMC article.
-
Integrin αDβ2 (CD11d/CD18) mediates experimental malaria-associated acute respiratory distress syndrome (MA-ARDS).Malar J. 2016 Jul 30;15(1):393. doi: 10.1186/s12936-016-1447-7. Malar J. 2016. PMID: 27473068 Free PMC article.
-
β2 Integrin CD11d/CD18: From Expression to an Emerging Role in Staged Leukocyte Migration.Front Immunol. 2021 Nov 8;12:775447. doi: 10.3389/fimmu.2021.775447. eCollection 2021. Front Immunol. 2021. PMID: 34858434 Free PMC article. Review.
-
Structural basis of blocking integrin activation and deactivation for anti-inflammation.J Biomed Sci. 2015 Jul 8;22(1):51. doi: 10.1186/s12929-015-0159-6. J Biomed Sci. 2015. PMID: 26152212 Free PMC article. Review.
Cited by
-
Humanized anti-CD11d monoclonal antibodies suitable for basic research and therapeutic applications.Antib Ther. 2024 Dec 16;8(1):26-39. doi: 10.1093/abt/tbae031. eCollection 2025 Jan. Antib Ther. 2024. PMID: 39839909 Free PMC article.
-
Multifaceted, unique role of CD11c in leukocyte biology.Front Immunol. 2025 Mar 4;16:1556992. doi: 10.3389/fimmu.2025.1556992. eCollection 2025. Front Immunol. 2025. PMID: 40103815 Free PMC article. Review.
-
Structural Immunology of Complement Receptors 3 and 4.Front Immunol. 2018 Nov 26;9:2716. doi: 10.3389/fimmu.2018.02716. eCollection 2018. Front Immunol. 2018. PMID: 30534123 Free PMC article. Review.
-
Mac-1 regulates disease stage-specific immunosuppression via the nitric oxide pathway in autoimmune disease.Sci Adv. 2025 May 9;11(19):eads3728. doi: 10.1126/sciadv.ads3728. Epub 2025 May 9. Sci Adv. 2025. PMID: 40344054 Free PMC article.
-
αDβ2 as a novel target of experimental polymicrobial sepsis.Front Immunol. 2022 Nov 18;13:1059996. doi: 10.3389/fimmu.2022.1059996. eCollection 2022. Front Immunol. 2022. PMID: 36466931 Free PMC article.
References
-
- Hynes RO (2002) Integrins: bidirectional, allosteric signaling machines. Cell 110:673–687. - PubMed
-
- Harris ES, McIntyre TM, Prescott SM, Zimmerman GA (2000) The leukocyte integrins. J Biol Chem 275:23409–23412. - PubMed
-
- Evans R, Patzak I, Svensson L, De Filippo K, Jones K, et al. (2009) Integrins in immunity. J Cell Sci 122:215–225. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
