

Length-dependent anisotropic scaling of spindle shape
- PMID: 25416062
- PMCID: PMC4265759
- DOI: 10.1242/bio.201410363
Length-dependent anisotropic scaling of spindle shape
Abstract
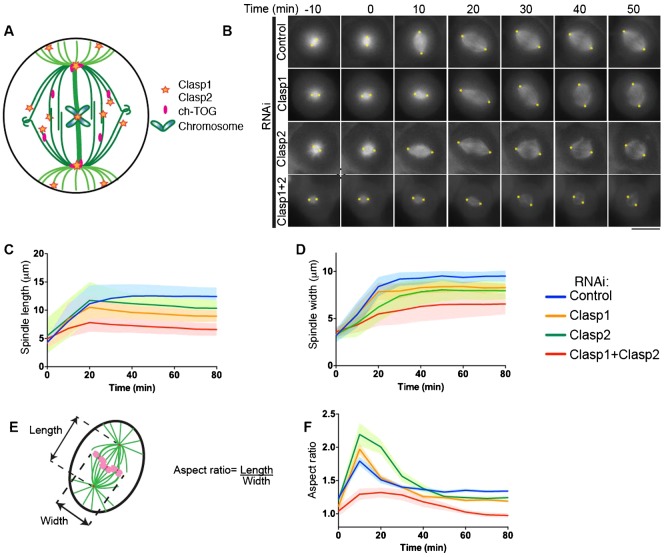
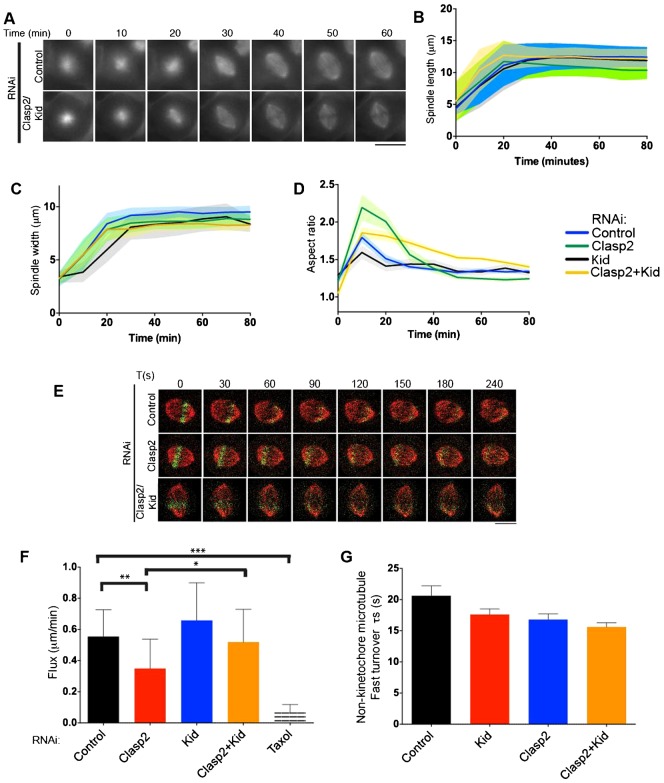
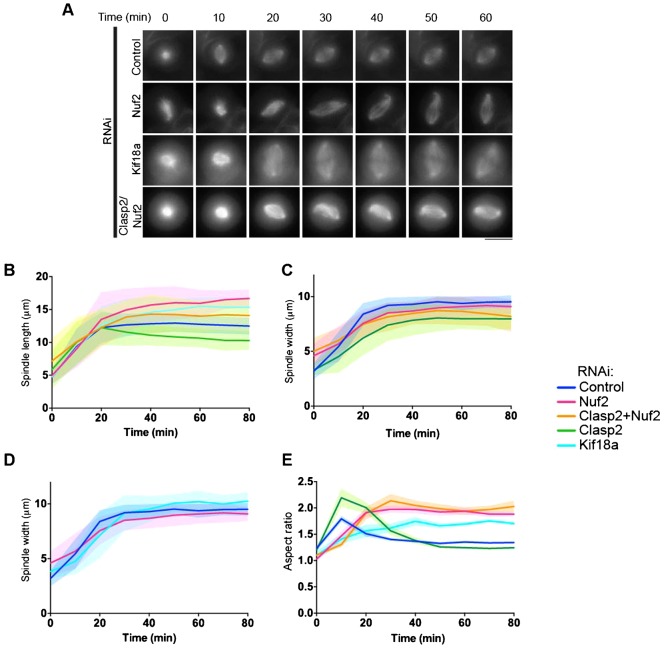
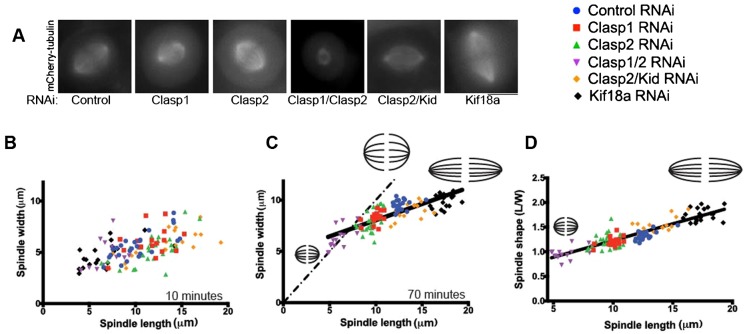
Spindle length varies dramatically across species and during early development to segregate chromosomes optimally. Both intrinsic factors, such as regulatory molecules, and extrinsic factors, such as cytoplasmic volume, determine spindle length scaling. However, the properties that govern spindle shape and whether these features can be modulated remain unknown. Here, we analyzed quantitatively how the molecular players which regulate microtubule dynamics control the kinetics of spindle formation and shape. We find that, in absence of Clasp1 and Clasp2, spindle assembly is biphasic due to unopposed inward pulling forces from the kinetochore-fibers and that kinetochore-fibers also alter spindle geometry. We demonstrate that spindle shape scaling is independent of the nature of the molecules that regulate dynamic microtubule properties, but is dependent on the steady-state metaphase spindle length. The shape of the spindle scales anisotropically with increasing length. Our results suggest that intrinsic mechanisms control the shape of the spindle to ensure the efficient capture and alignment of chromosomes independently of spindle length.
Keywords: Clasp; K-fiber; Microtubules; Mitosis; Spindle.
© 2014. Published by The Company of Biologists Ltd.
Conflict of interest statement
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
