

Evolving gene regulatory networks into cellular networks guiding adaptive behavior: an outline how single cells could have evolved into a centralized neurosensory system
- PMID: 25416504
- PMCID: PMC4286516
- DOI: 10.1007/s00441-014-2043-1
Evolving gene regulatory networks into cellular networks guiding adaptive behavior: an outline how single cells could have evolved into a centralized neurosensory system
Abstract
Understanding the evolution of the neurosensory system of man, able to reflect on its own origin, is one of the major goals of comparative neurobiology. Details of the origin of neurosensory cells, their aggregation into central nervous systems and associated sensory organs and their localized patterning leading to remarkably different cell types aggregated into variably sized parts of the central nervous system have begun to emerge. Insights at the cellular and molecular level have begun to shed some light on the evolution of neurosensory cells, partially covered in this review. Molecular evidence suggests that high mobility group (HMG) proteins of pre-metazoans evolved into the definitive Sox [SRY (sex determining region Y)-box] genes used for neurosensory precursor specification in metazoans. Likewise, pre-metazoan basic helix-loop-helix (bHLH) genes evolved in metazoans into the group A bHLH genes dedicated to neurosensory differentiation in bilaterians. Available evidence suggests that the Sox and bHLH genes evolved a cross-regulatory network able to synchronize expansion of precursor populations and their subsequent differentiation into novel parts of the brain or sensory organs. Molecular evidence suggests metazoans evolved patterning gene networks early, which were not dedicated to neuronal development. Only later in evolution were these patterning gene networks tied into the increasing complexity of diffusible factors, many of which were already present in pre-metazoans, to drive local patterning events. It appears that the evolving molecular basis of neurosensory cell development may have led, in interaction with differentially expressed patterning genes, to local network modifications guiding unique specializations of neurosensory cells into sensory organs and various areas of the central nervous system.
Figures
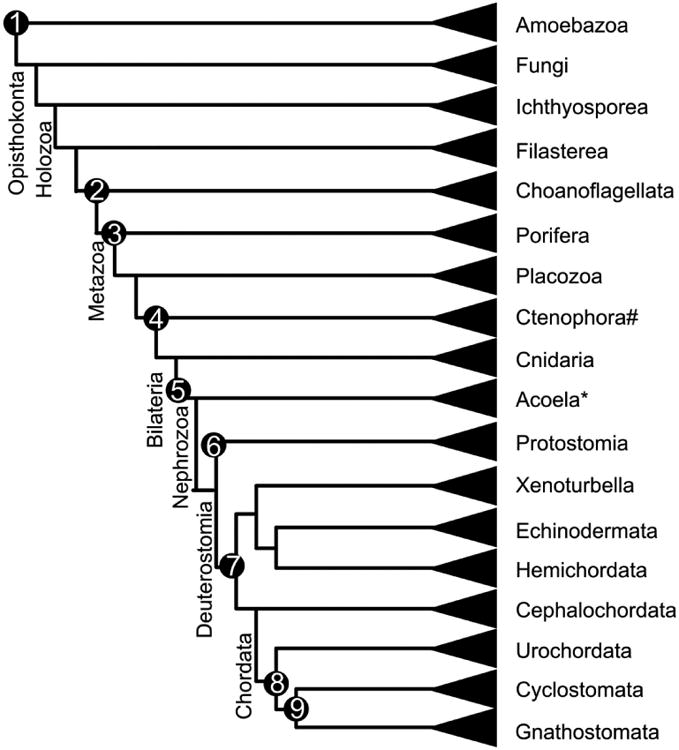
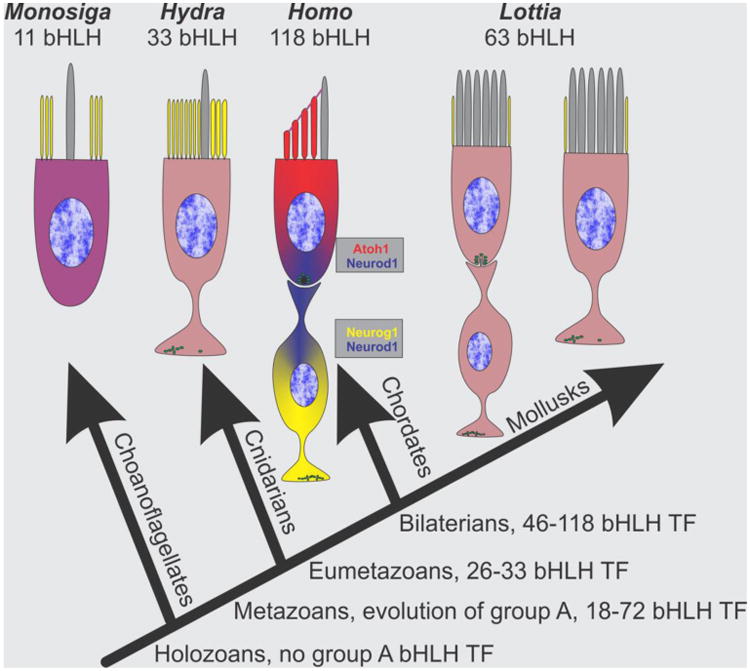
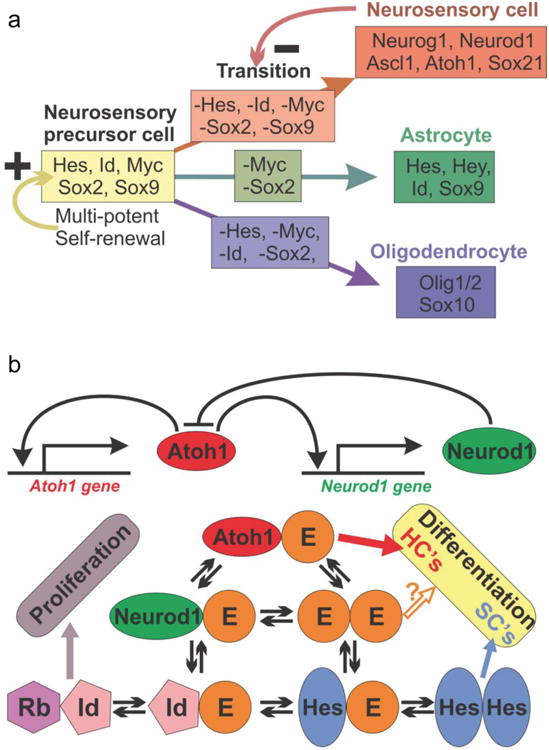
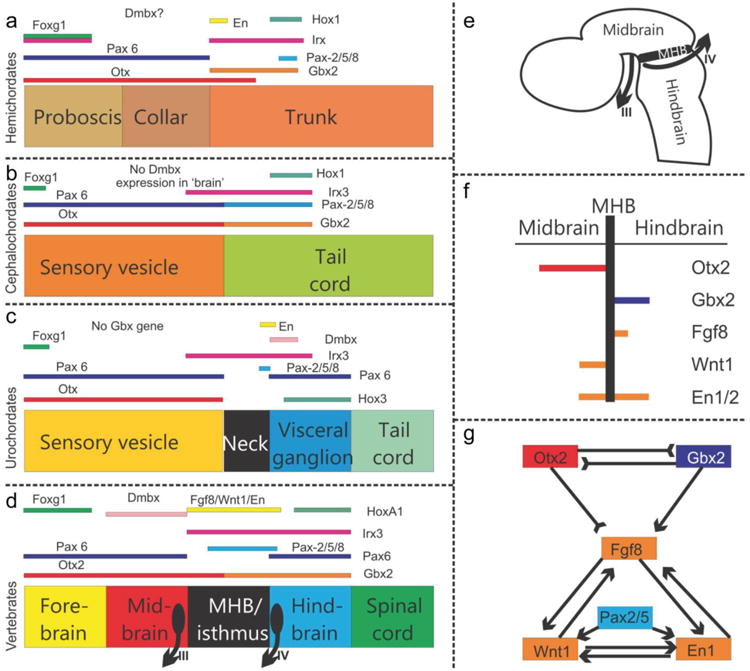
References
-
- Albert B, Colleran C, Leger-Silvestre I, Berger AB, Dez C, Normand C, Perez-Fernandez J, McStay B, Gadal O. Structure-function analysis of Hmo1 unveils an ancestral organization of HMG-Box factors involved in ribosomal DNA transcription from yeast to human. Nucleic acids research. 2013;41:10135–10149. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
