

NCoR1 and SMRT play unique roles in thyroid hormone action in vivo
- PMID: 25421714
- PMCID: PMC4285426
- DOI: 10.1128/MCB.01208-14
NCoR1 and SMRT play unique roles in thyroid hormone action in vivo
Abstract
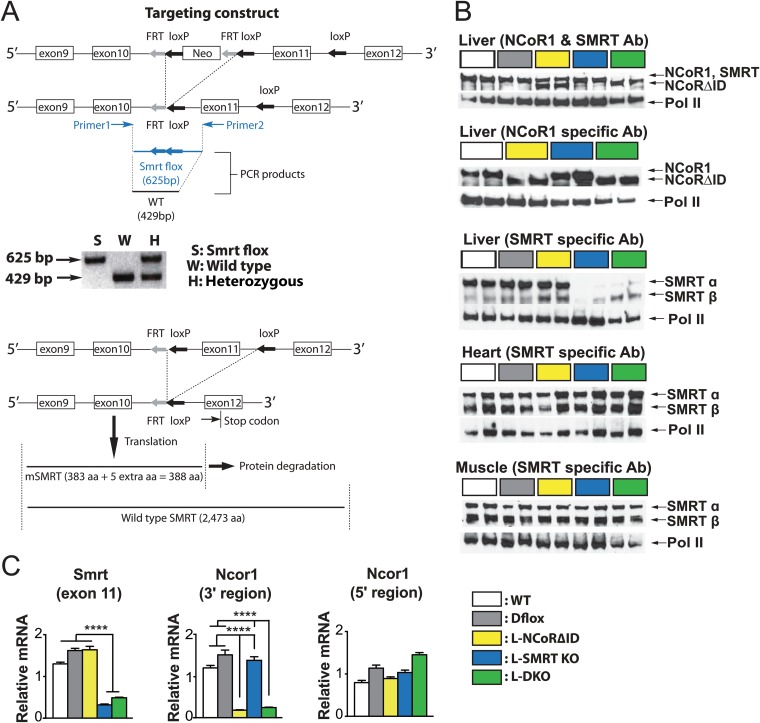
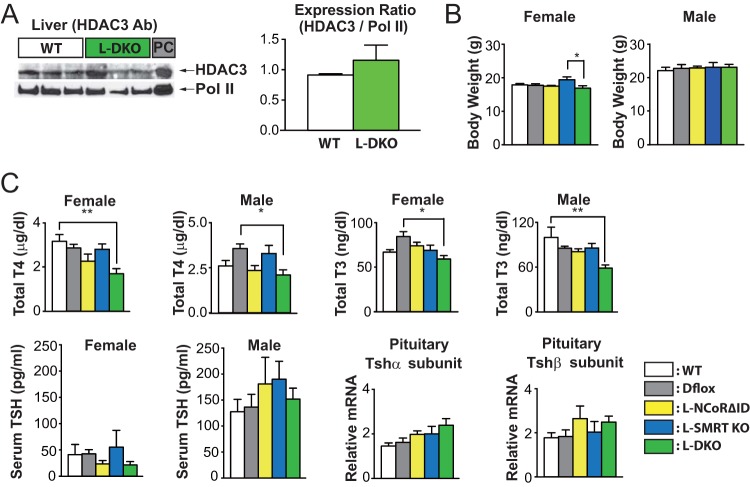
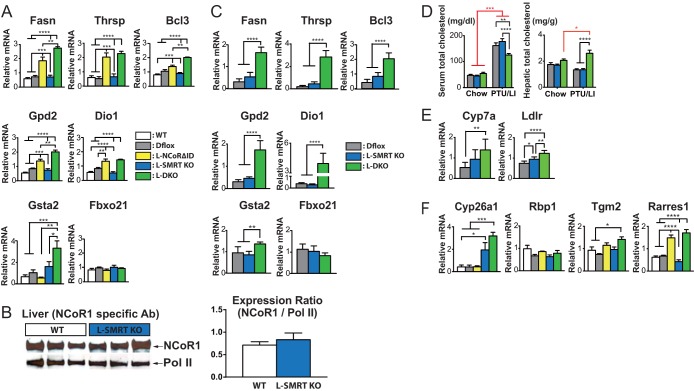
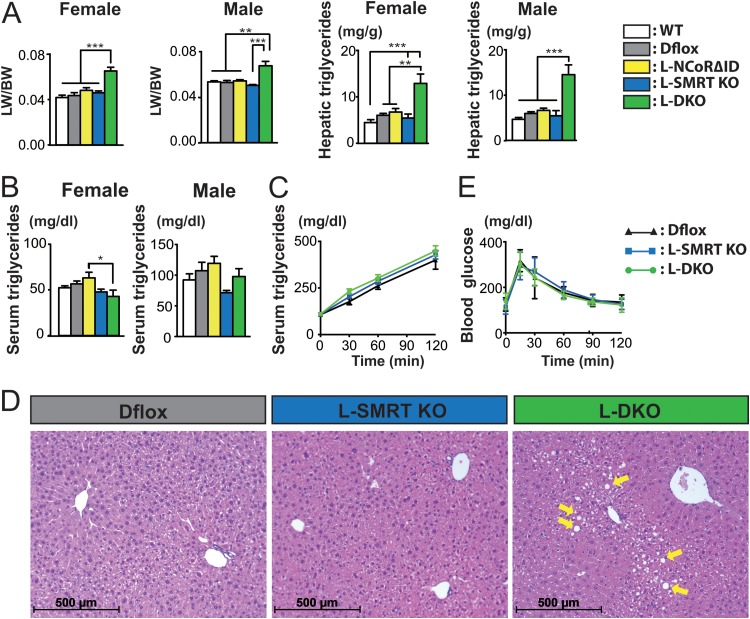
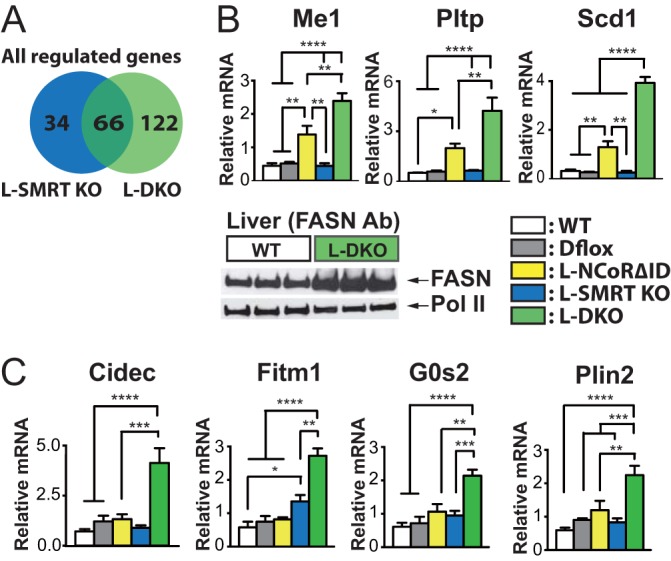
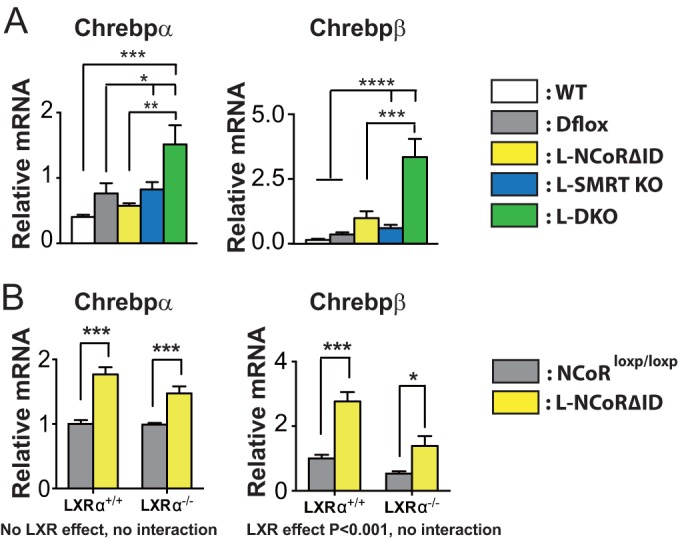
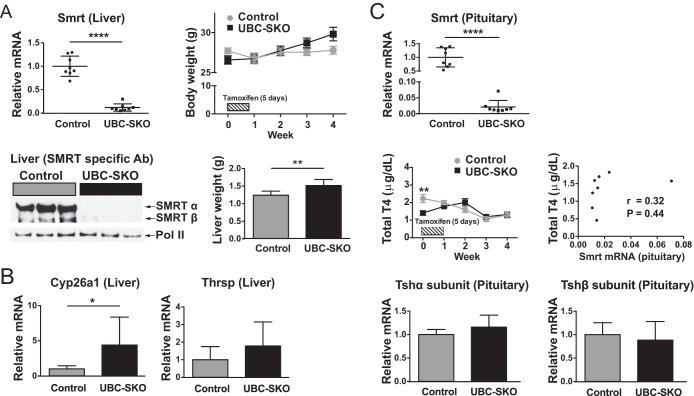
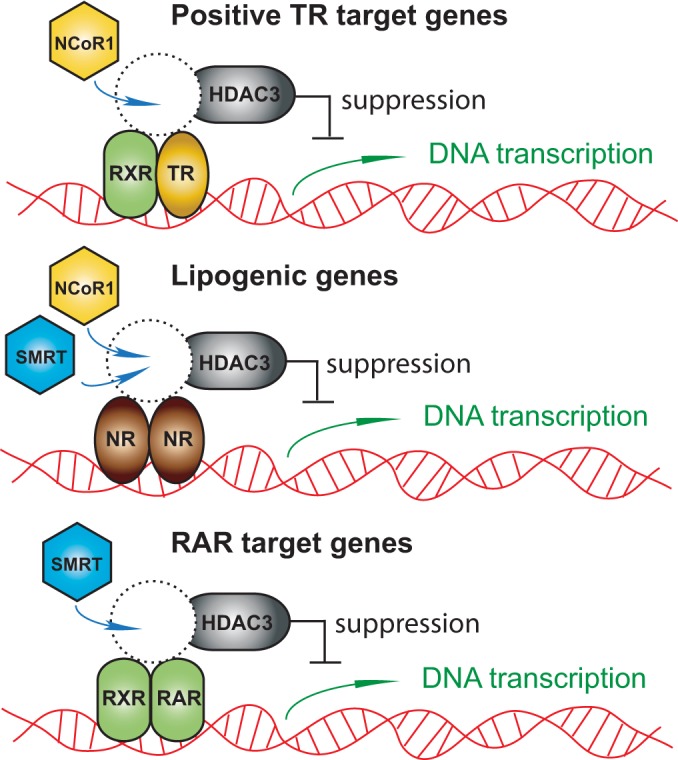
NCoR1 (nuclear receptor corepressor) and SMRT (silencing mediator of retinoid and thyroid hormone receptors; NCoR2) are well-recognized coregulators of nuclear receptor (NR) action. However, their unique roles in the regulation of thyroid hormone (TH) signaling in specific cell types have not been determined. To accomplish this we generated mice that lacked function of either NCoR1, SMRT, or both in the liver only and additionally a global SMRT knockout model. Despite both corepressors being present in the liver, deletion of SMRT in either euthyroid or hypothyroid animals had little effect on TH signaling. In contrast, disruption of NCoR1 action confirmed that NCoR1 is the principal mediator of TH sensitivity in vivo. Similarly, global disruption of SMRT, unlike the global disruption of NCoR1, did not affect TH levels. While SMRT played little role in TH-regulated pathways, when disrupted in combination with NCoR1, it greatly accentuated the synthesis and storage of hepatic lipid. Taken together, these data demonstrate that corepressor specificity exists in vivo and that NCoR1 is the principal regulator of TH action. However, both corepressors collaborate to control hepatic lipid content, which likely reflects their cooperative activity in regulating the action of multiple NRs including the TH receptor (TR).
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Nuclear Receptor CoRepressors, NCOR1 and SMRT, are required for maintaining systemic metabolic homeostasis.Mol Metab. 2021 Nov;53:101315. doi: 10.1016/j.molmet.2021.101315. Epub 2021 Aug 12. Mol Metab. 2021. PMID: 34390859 Free PMC article.
-
GR SUMOylation and formation of an SUMO-SMRT/NCoR1-HDAC3 repressing complex is mandatory for GC-induced IR nGRE-mediated transrepression.Proc Natl Acad Sci U S A. 2016 Feb 2;113(5):E626-34. doi: 10.1073/pnas.1522821113. Epub 2015 Dec 28. Proc Natl Acad Sci U S A. 2016. PMID: 26712002 Free PMC article.
-
Emerging roles of the corepressors NCoR1 and SMRT in homeostasis.Genes Dev. 2013 Apr 15;27(8):819-35. doi: 10.1101/gad.214023.113. Genes Dev. 2013. PMID: 23630073 Free PMC article. Review.
-
Nuclear corepressor SMRT is a strong regulator of body weight independently of its ability to regulate thyroid hormone action.PLoS One. 2019 Aug 12;14(8):e0220717. doi: 10.1371/journal.pone.0220717. eCollection 2019. PLoS One. 2019. PMID: 31404087 Free PMC article.
-
Transcriptional Cofactors for Thyroid Hormone Receptors.Endocrinology. 2025 Jan 6;166(2):bqae164. doi: 10.1210/endocr/bqae164. Endocrinology. 2025. PMID: 39679543 Review.
Cited by
-
HDAC3 Is a Master Regulator of mTEC Development.Cell Rep. 2016 Apr 19;15(3):651-665. doi: 10.1016/j.celrep.2016.03.048. Epub 2016 Apr 7. Cell Rep. 2016. PMID: 27068467 Free PMC article.
-
Thyroid hormone: sex-dependent role in nervous system regulation and disease.Biol Sex Differ. 2021 Mar 8;12(1):25. doi: 10.1186/s13293-021-00367-2. Biol Sex Differ. 2021. PMID: 33685490 Free PMC article. Review.
-
Critical Period Regulation by Thyroid Hormones: Potential Mechanisms and Sex-Specific Aspects.Front Mol Neurosci. 2019 Apr 5;12:77. doi: 10.3389/fnmol.2019.00077. eCollection 2019. Front Mol Neurosci. 2019. PMID: 31024251 Free PMC article.
-
Thyroid Hormones and Co-workers: An Overview.Methods Mol Biol. 2025;2876:3-16. doi: 10.1007/978-1-0716-4252-8_1. Methods Mol Biol. 2025. PMID: 39579305 Review.
-
HDAC3 ensures stepwise epidermal stratification via NCoR/SMRT-reliant mechanisms independent of its histone deacetylase activity.Genes Dev. 2020 Jul 1;34(13-14):973-988. doi: 10.1101/gad.333674.119. Epub 2020 May 28. Genes Dev. 2020. PMID: 32467224 Free PMC article.
References
-
- Yen PM. 2001. Physiological and molecular basis of thyroid hormone action. Physiol Rev 81:1097–1142. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials