

The RapP-PhrP quorum-sensing system of Bacillus subtilis strain NCIB3610 affects biofilm formation through multiple targets, due to an atypical signal-insensitive allele of RapP
- PMID: 25422306
- PMCID: PMC4285980
- DOI: 10.1128/JB.02382-14
The RapP-PhrP quorum-sensing system of Bacillus subtilis strain NCIB3610 affects biofilm formation through multiple targets, due to an atypical signal-insensitive allele of RapP
Abstract
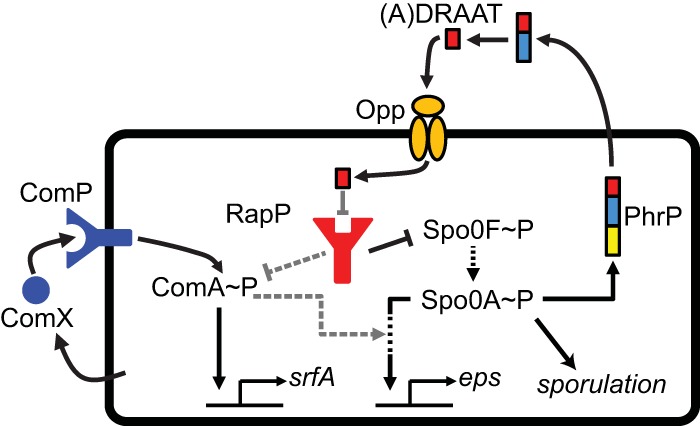
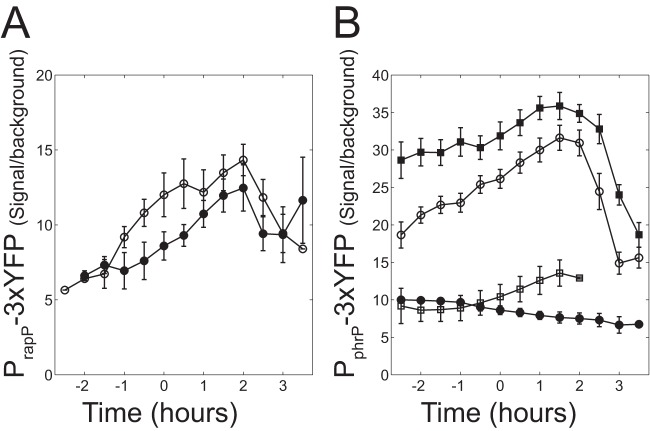
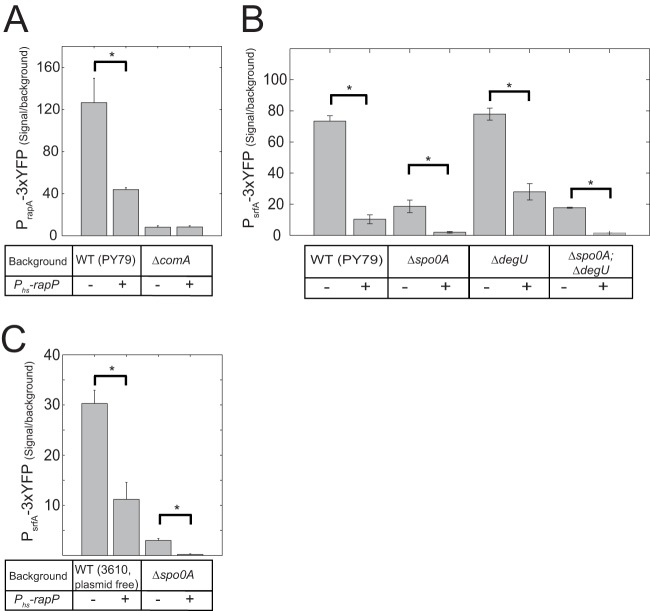
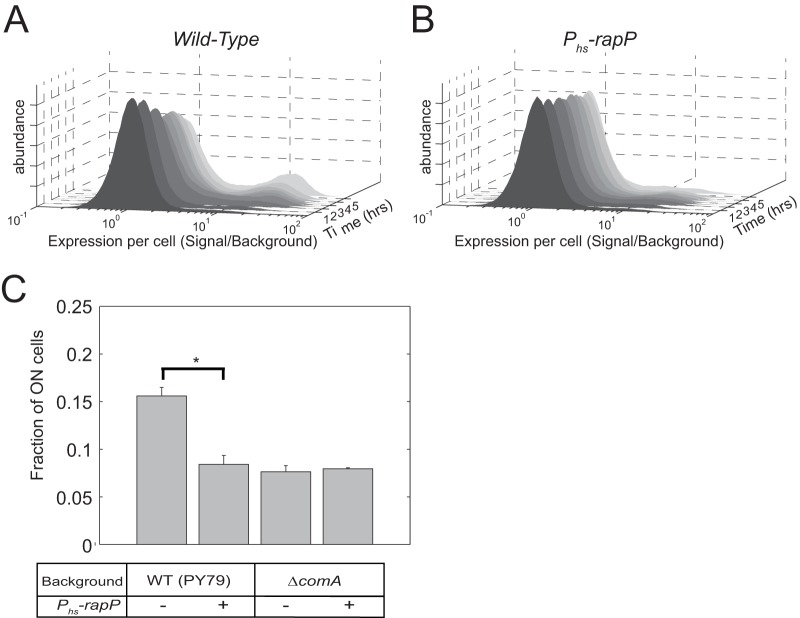
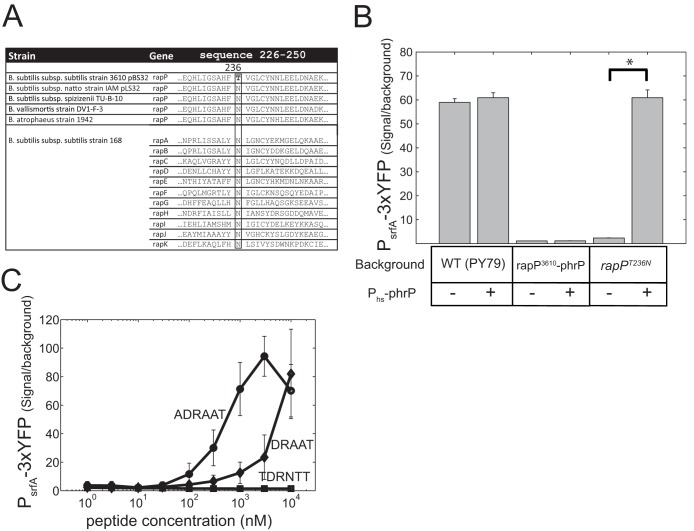
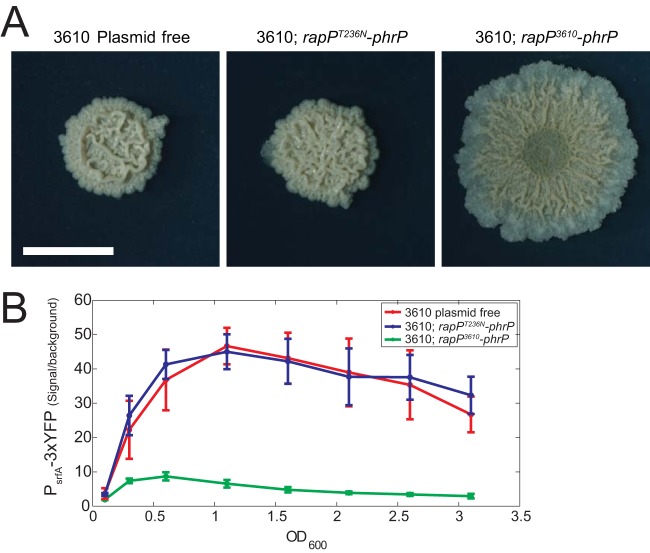
The genome of Bacillus subtilis 168 encodes eight rap-phr quorum-sensing pairs. Rap proteins of all characterized Rap-Phr pairs inhibit the function of one or several important response regulators: ComA, Spo0F, or DegU. This inhibition is relieved upon binding of the peptide encoded by the cognate phr gene. Bacillus subtilis strain NCIB3610, the biofilm-proficient ancestor of strain 168, encodes, in addition, the rapP-phrP pair on the plasmid pBS32. RapP was shown to dephosphorylate Spo0F and to regulate biofilm formation, but unlike other Rap-Phr pairs, RapP does not interact with PhrP. In this work we extend the analysis of the RapP pathway by reexamining its transcriptional regulation, its effect on downstream targets, and its interaction with PhrP. At the transcriptional level, we show that rapP and phrP regulation is similar to that of other rap-phr pairs. We further find that RapP has an Spo0F-independent negative effect on biofilm-related genes, which is mediated by the response regulator ComA. Finally, we find that the insensitivity of RapP to PhrP is due to a substitution of a highly conserved residue in the peptide binding domain of the rapP allele of strain NCIB3610. Reversing this substitution to the consensus amino acid restores the PhrP dependence of RapP activity and eliminates the effects of the rapP-phrP locus on ComA activity and biofilm formation. Taken together, our results suggest that RapP strongly represses biofilm formation through multiple targets and that PhrP does not counteract RapP due to a rare mutation in rapP.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
The Large pBS32/pLS32 Plasmid of Ancestral Bacillus subtilis.J Bacteriol. 2020 Aug 25;202(18):e00290-20. doi: 10.1128/JB.00290-20. Print 2020 Aug 25. J Bacteriol. 2020. PMID: 32601071 Free PMC article. Review.
-
A plasmid-encoded phosphatase regulates Bacillus subtilis biofilm architecture, sporulation, and genetic competence.J Bacteriol. 2013 May;195(10):2437-48. doi: 10.1128/JB.02030-12. Epub 2013 Mar 22. J Bacteriol. 2013. PMID: 23524609 Free PMC article.
-
Novel mechanisms of controlling the activities of the transcription factors Spo0A and ComA by the plasmid-encoded quorum sensing regulators Rap60-Phr60 in Bacillus subtilis.Mol Microbiol. 2015 Apr;96(2):325-48. doi: 10.1111/mmi.12939. Epub 2015 Feb 18. Mol Microbiol. 2015. PMID: 25598361 Free PMC article.
-
A complex path for domestication of B. subtilis sociality.Curr Genet. 2015 Nov;61(4):493-6. doi: 10.1007/s00294-015-0479-9. Epub 2015 Feb 14. Curr Genet. 2015. PMID: 25680358 Review.
-
Rap Protein Paralogs of Bacillus thuringiensis: a Multifunctional and Redundant Regulatory Repertoire for the Control of Collective Functions.J Bacteriol. 2020 Feb 25;202(6):e00747-19. doi: 10.1128/JB.00747-19. Print 2020 Feb 25. J Bacteriol. 2020. PMID: 31871034 Free PMC article.
Cited by
-
The Exo-Polysaccharide Component of Extracellular Matrix is Essential for the Viscoelastic Properties of Bacillus subtilis Biofilms.Int J Mol Sci. 2020 Sep 15;21(18):6755. doi: 10.3390/ijms21186755. Int J Mol Sci. 2020. PMID: 32942569 Free PMC article.
-
RnhP is a plasmid-borne RNase HI that contributes to genome maintenance in the ancestral strain Bacillus subtilis NCIB 3610.Mol Microbiol. 2021 Jan;115(1):99-115. doi: 10.1111/mmi.14601. Epub 2020 Sep 25. Mol Microbiol. 2021. PMID: 32896031 Free PMC article.
-
Molecular and cellular factors control signal transduction via switchable allosteric modulator proteins (SAMPs).BMC Syst Biol. 2016 Apr 27;10:35. doi: 10.1186/s12918-016-0274-3. BMC Syst Biol. 2016. PMID: 27122155 Free PMC article.
-
The putative role of the epipeptide EpeX in Bacillus subtilis intra-species competition.Microbiology (Reading). 2023 Jun;169(6):001344. doi: 10.1099/mic.0.001344. Microbiology (Reading). 2023. PMID: 37289492 Free PMC article.
-
The Large pBS32/pLS32 Plasmid of Ancestral Bacillus subtilis.J Bacteriol. 2020 Aug 25;202(18):e00290-20. doi: 10.1128/JB.00290-20. Print 2020 Aug 25. J Bacteriol. 2020. PMID: 32601071 Free PMC article. Review.
References
-
- Comella N, Grossman AD. 2005. Conservation of genes and processes controlled by the quorum response in bacteria: characterization of genes controlled by the quorum-sensing transcription factor ComA in Bacillus subtilis. Mol Microbiol 57:1159–1174. doi:10.1111/j.1365-2958.2005.04749.x. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
