

Review
doi: 10.1083/jcb.201407015.
Shaping the intestinal brush border
Affiliations
- PMID: 25422372
- PMCID: PMC4242837
- DOI: 10.1083/jcb.201407015
Item in Clipboard
Review
Shaping the intestinal brush border
J Cell Biol.
.
Abstract
Epithelial cells from diverse tissues, including the enterocytes that line the intestinal tract, remodel their apical surface during differentiation to form a brush border: an array of actin-supported membrane protrusions known as microvilli that increases the functional capacity of the tissue. Although our understanding of how epithelial cells assemble, stabilize, and organize apical microvilli is still developing, investigations of the biochemical and physical underpinnings of these processes suggest that cells coordinate cytoskeletal remodeling, membrane-cytoskeleton cross-linking, and extracellular adhesion to shape the apical brush border domain.
© 2014 Crawley et al.
Figures
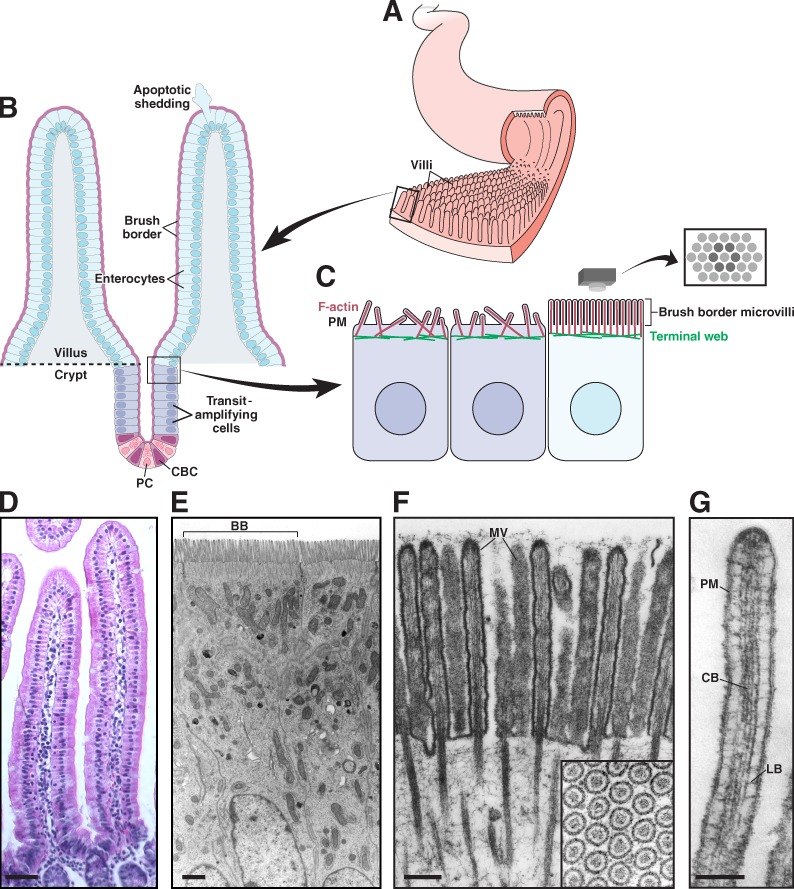
Functional architecture of the intestinal epithelium. (A) The small intestinal epithelium is characterized by ubiquitous small folds of tissue known as villi. (B) Enterocytes, the most abundant cell type lining the villus, are generated in a stem cell niche composed of crypt base columnar (CBC) cells and flanking Paneth cells (PC), found in crypts near the base of each villus. CBC cells undergo asymmetric cell division, resulting in a new stem cell plus a committed daughter cell. Daughter cells undergo further division in the transit-amplifying region before differentiating into enterocytes and exiting the crypt. Enterocytes migrate up the crypt–villus axis over a period of 2–3 d. Once at the tip, cells undergo apoptosis and are extruded from the epithelium. (C) Apical surface organization of enterocytes before and after differentiation; brush border microvilli on the surface of differentiated cells are packed in tight, hexagonal arrays. (D) Villi from mouse small intestine. (E) Enterocytes from mouse small intestine; brush border (BB) from a single cell is indicated. (F) Brush border microvilli (MV) from chicken small intestine. Reproduced from Mooseker and Tilney (1975). The inset shows hexagonally packed microvilli in cross section. (G) A single microvillus. Plasma membrane (PM) is linked to the core bundle (CB) by lateral bridges (LB) that are formed at least in part by myo1a. Reproduced from Mooseker and Tilney (1975). Bars: (D) 50 µm; (E) 1 µm; (F) 0.2 µm; (G) 0.1 µm.
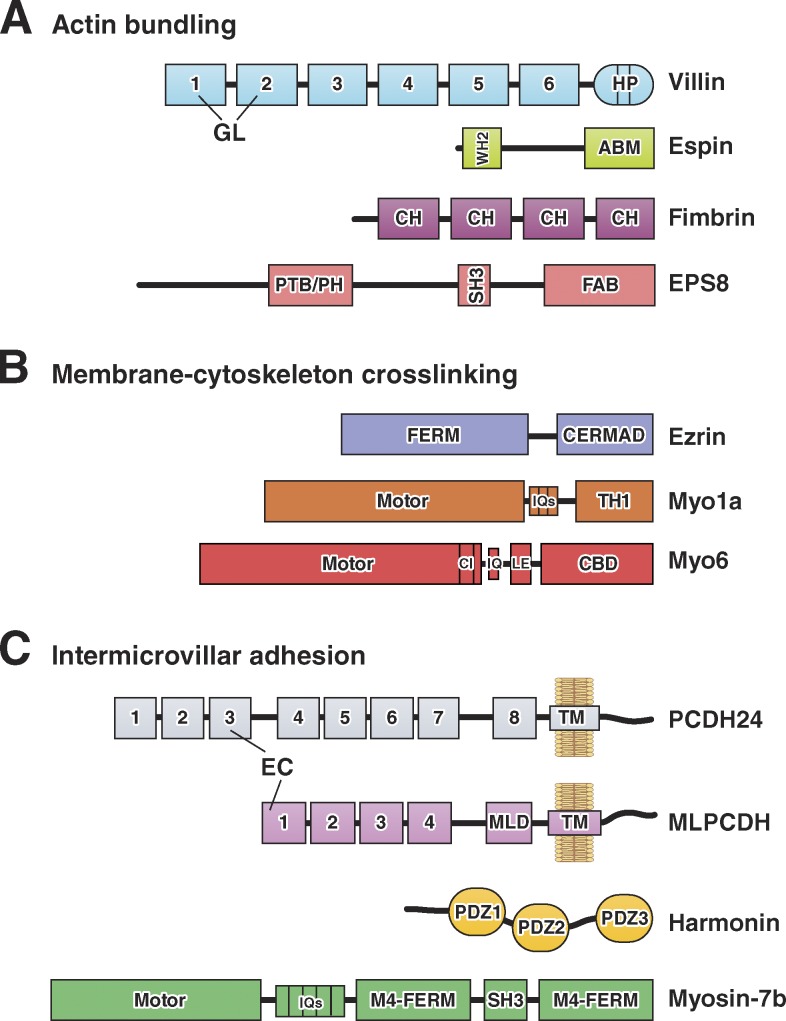
Domain organization of F-actin bundling, membrane–cytoskeleton cross-linking, and intermicrovillar adhesion molecules. (A) Actin-bundling. Villin is composed of multiple gelsolin-like (GL) domains and a C-terminal headpiece (HP) domain. Small espin contains a G-actin–binding Wiskott-Aldrich homology 2 (WH2) domain and an actin-bundling motif (ABM). Fimbrin contains tandem copies of a conserved calponin homology (CH) motif that binds actin. EPS8 contains phosphotyrosine-binding (PTB), pleckstrin homology (PH), SH3, and F-actin bundling (FAB) domains. (B) Membrane–cytoskeleton cross-linking. Ezrin contains a FERM domain and an ERM-association domain (C-ERMAD). The myo1a motor domain is linked to CaM-binding IQ motifs and a membrane-binding TH1 domain. The myo6 motor domain is linked to a converter insert (CI), an IQ motif, a lever arm extension (LE), and a cargo-binding domain (CBD). (C) Intermicrovillar adhesion. PCDH24 and MLPCDH contain multiple extracellular cadherin (EC) repeats; MLPCDH also contains a juxtamembrane mucin-like domain (MLD). Harmonin-a is composed of three PDZ domains. Myosin-7b contains an N-terminal motor domain, CaM-binding IQ motifs, and a tail composed of two MyTH4-FERM domains with an intervening SH3 domain.
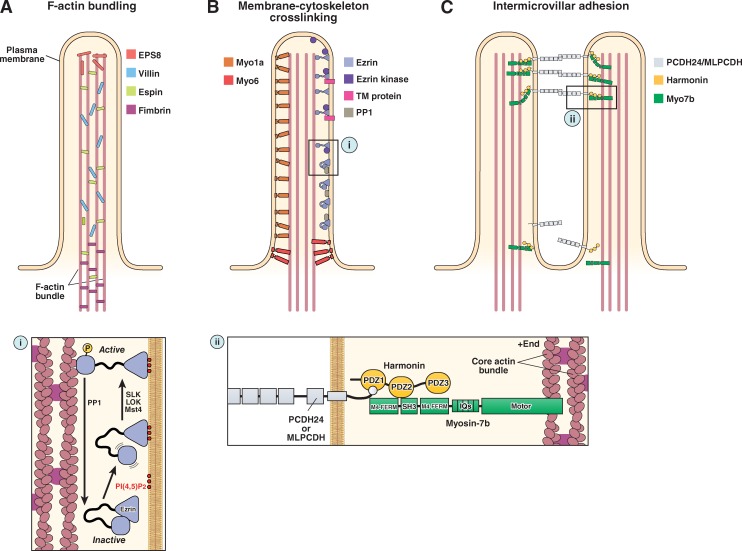
The molecular machinery of microvillar protrusion, stabilization, and organization. (A) Bundling of actin filaments in the microvillar actin core is performed through the collective and potentially redundant function of villin, espin, fimbrin, and EPS8. (B) Membrane–cytoskeleton cross-linking plays an important role in microvillar stabilization and is mediated by myo1a, myo6, and the active form of ezrin (i). (C) Extracellular adhesion between the distal tips of microvilli (i.e., intermicrovillar adhesion) is used to optimize the packing of these protrusions during brush border assembly. Intermicrovillar adhesion is mediated by a trans-heterophilic complex of PCDH24 and MLPCDH. Distal tip targeting of microvillar protocadherins requires interactions with harmonin and, potentially, myosin-7b (ii). See main text for details on the function of the other proteins depicted in the figure.
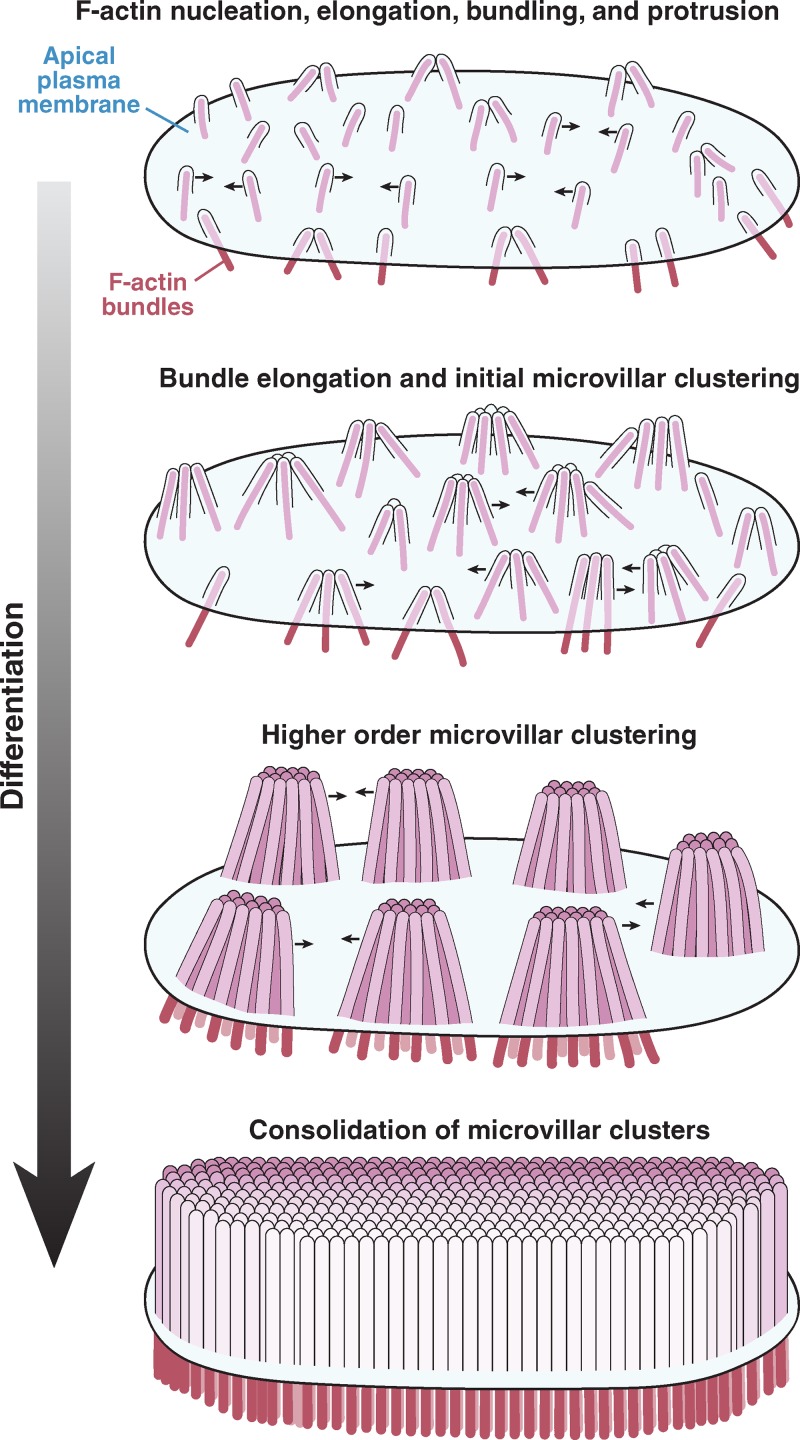
Remodeling of the apical surface during enterocyte differentiation. Microvilli initially appear as small membrane buds on the apical surface, formed by the polymerization and bundling of actin filaments beneath the membrane. From an early time point, these protrusions self-organize into distinct clusters, a process that is mediated by protocadherin-based adhesion between their distal tips. Distal tip adhesion promotes the highly ordered packing of microvilli, as well as uniformity in microvillar length. As maturation continues, clusters grow (in terms of number of microvilli incorporated) and continue to consolidate, ultimately resulting in a single large-scale cluster: a fully differentiated brush border.
References
-
- Ahmed Z.M., Goodyear R., Riazuddin S., Lagziel A., Legan P.K., Behra M., Burgess S.M., Lilley K.S., Wilcox E.R., Riazuddin S., et al. 2006. The tip-link antigen, a protein associated with the transduction complex of sensory hair cells, is protocadherin-15. J. Neurosci. 26:7022–7034 10.1523/JNEUROSCI.1163-06.2006 - DOI - PMC - PubMed
-
- Alagramam K.N., Goodyear R.J., Geng R., Furness D.N., van Aken A.F., Marcotti W., Kros C.J., and Richardson G.P.. 2011. Mutations in protocadherin 15 and cadherin 23 affect tip links and mechanotransduction in mammalian sensory hair cells. PLoS ONE. 6:e19183 10.1371/journal.pone.0019183 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
