

Potential role of orexin and sleep modulation in the pathogenesis of Alzheimer's disease
- PMID: 25422493
- PMCID: PMC4267230
- DOI: 10.1084/jem.20141788
Potential role of orexin and sleep modulation in the pathogenesis of Alzheimer's disease
Erratum in
- J Exp Med. 2015 Jan 12;212(1):121
Abstract
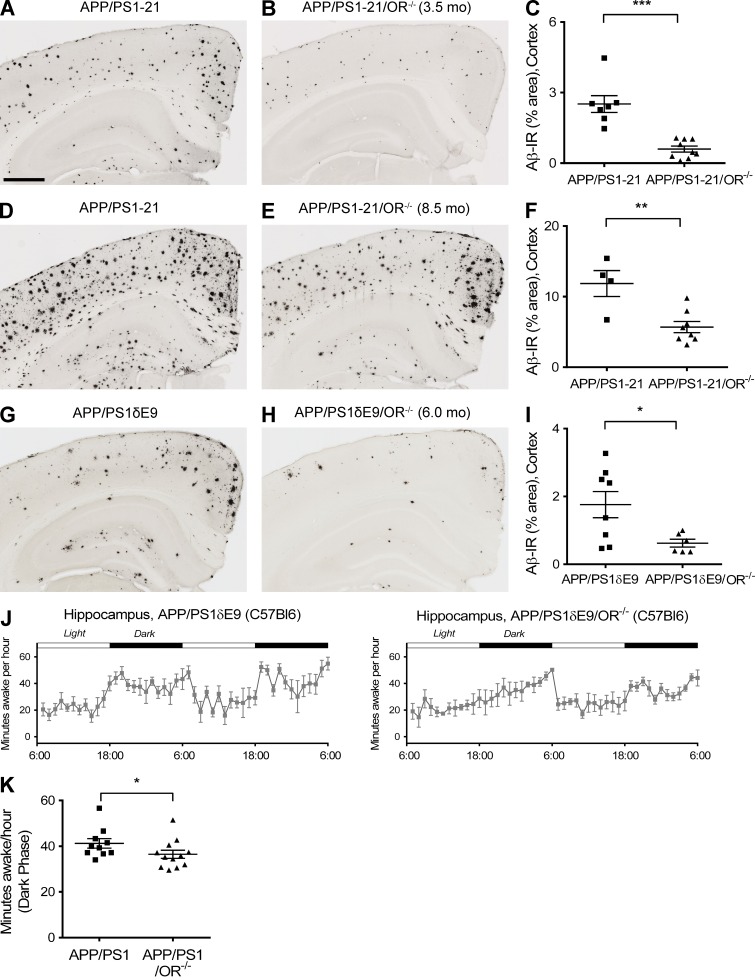
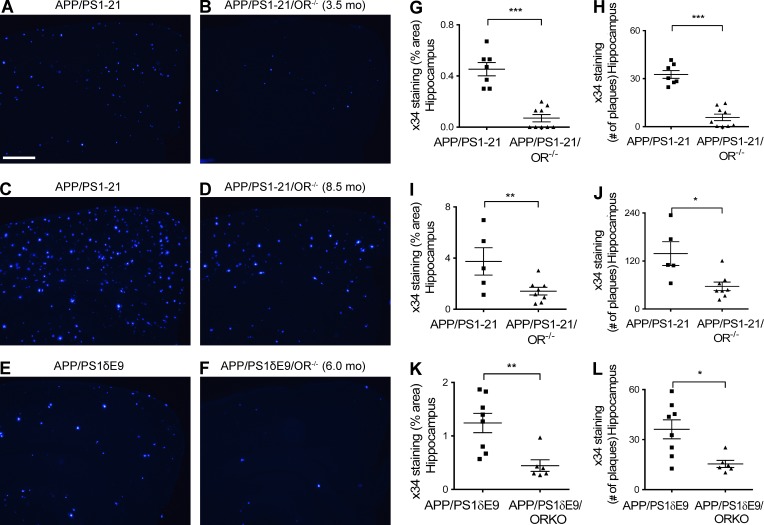
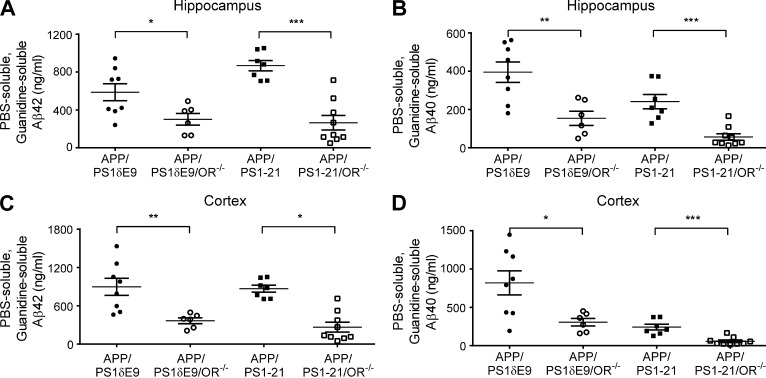
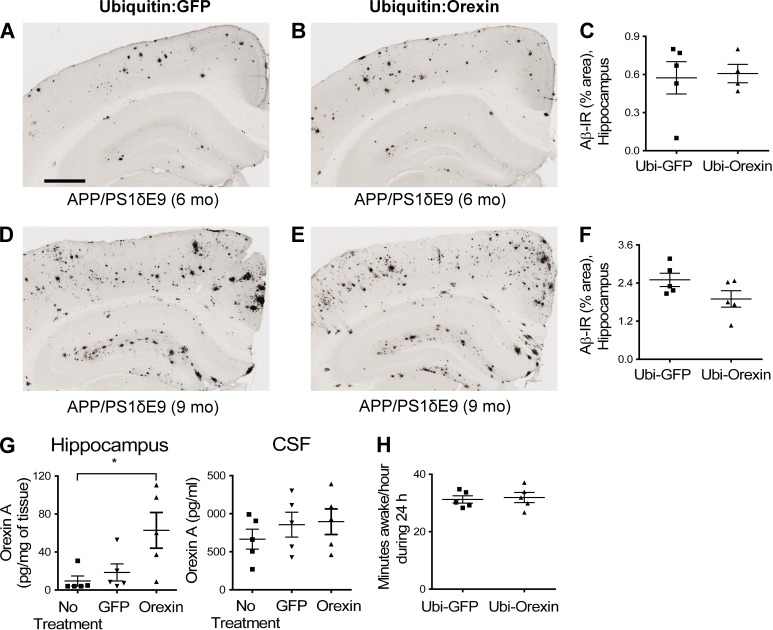
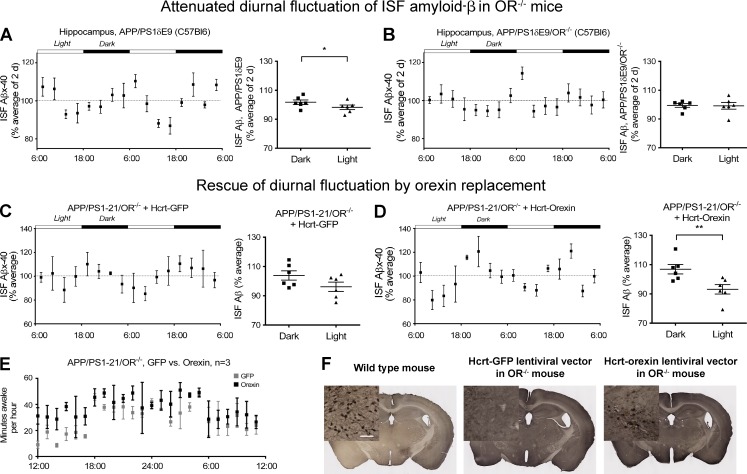
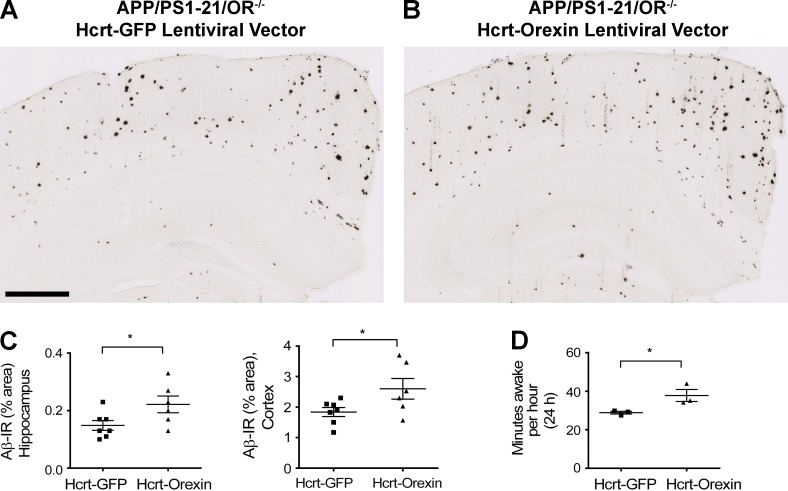
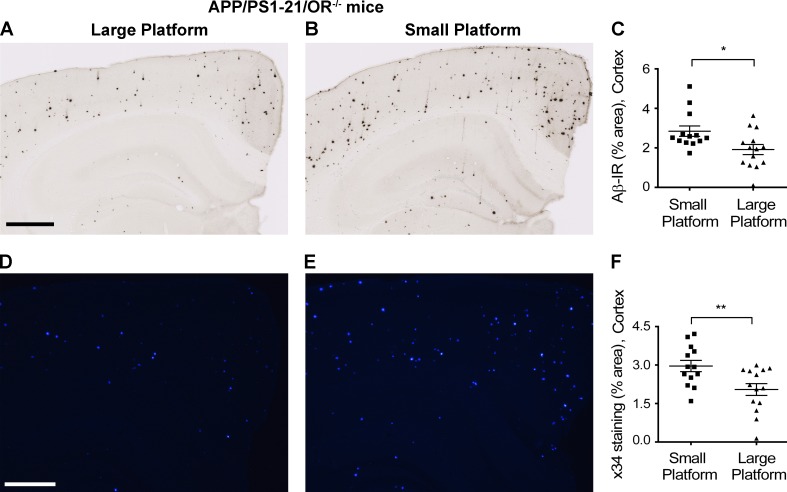
Age-related aggregation of amyloid-β (Aβ) is an upstream pathological event in Alzheimer's disease (AD) pathogenesis, and it disrupts the sleep-wake cycle. The amount of sleep declines with aging and to a greater extent in AD. Poor sleep quality and insufficient amounts of sleep have been noted in humans with preclinical evidence of AD. However, how the amount and quality of sleep affects Aβ aggregation is not yet well understood. Orexins (hypocretins) initiate and maintain wakefulness, and loss of orexin-producing neurons causes narcolepsy. We tried to determine whether orexin release or secondary changes in sleep via orexin modulation affect Aβ pathology. Amyloid precursor protein (APP)/Presenilin 1 (PS1) transgenic mice, in which the orexin gene is knocked out, showed a marked decrease in the amount of Aβ pathology in the brain with an increase in sleep time. Focal overexpression of orexin in the hippocampus in APP/PS1 mice did not alter the total amount of sleep/wakefulness and the amount of Aβ pathology. In contrast, sleep deprivation or increasing wakefulness by rescue of orexinergic neurons in APP/PS1 mice lacking orexin increased the amount of Aβ pathology in the brain. Collectively, modulation of orexin and its effects on sleep appear to modulate Aβ pathology in the brain.
© 2014 Roh et al.
Figures
Comment in
-
Sleeping our Alzheimer's risk away.J Exp Med. 2014 Dec 15;211(13):2484-5. doi: 10.1084/jem.21113insight4. J Exp Med. 2014. PMID: 25512582 Free PMC article. No abstract available.
References
-
- Cirrito J.R., May P.C., O’Dell M.A., Taylor J.W., Parsadanian M., Cramer J.W., Audia J.E., Nissen J.S., Bales K.R., Paul S.M., et al. . 2003. In vivo assessment of brain interstitial fluid with microdialysis reveals plaque-associated changes in amyloid-β metabolism and half-life. J. Neurosci. 23:8844–8853. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
