

Review
doi: 10.1021/cr500350x.
Epub 2014 Nov 26.
Post-translational modifications of histones that influence nucleosome dynamics
Affiliations
- PMID: 25424540
- PMCID: PMC4375056
- DOI: 10.1021/cr500350x
Item in Clipboard
Review
Post-translational modifications of histones that influence nucleosome dynamics
Chem Rev.
.
No abstract available
Figures
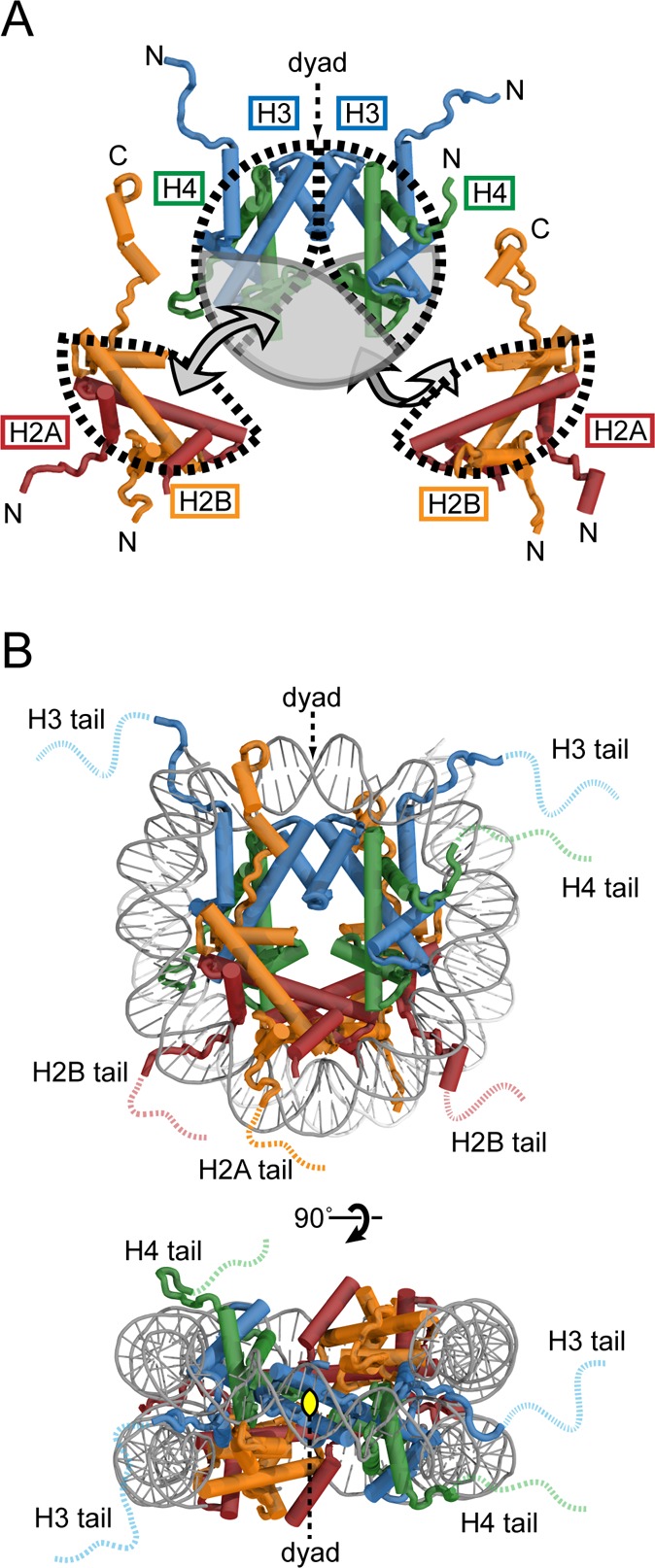
Overview of nucleosome architecture. (A) Illustration of H2A/H2B
and H3/H4 heterodimers and how they fit together to form the histone
octamer. (B) Face and top view of the nucleosome structure. For this
and all subsequent molecular representations of the nucleosome, the
high-resolution crystal structure (PDB code 1KX5) was used.
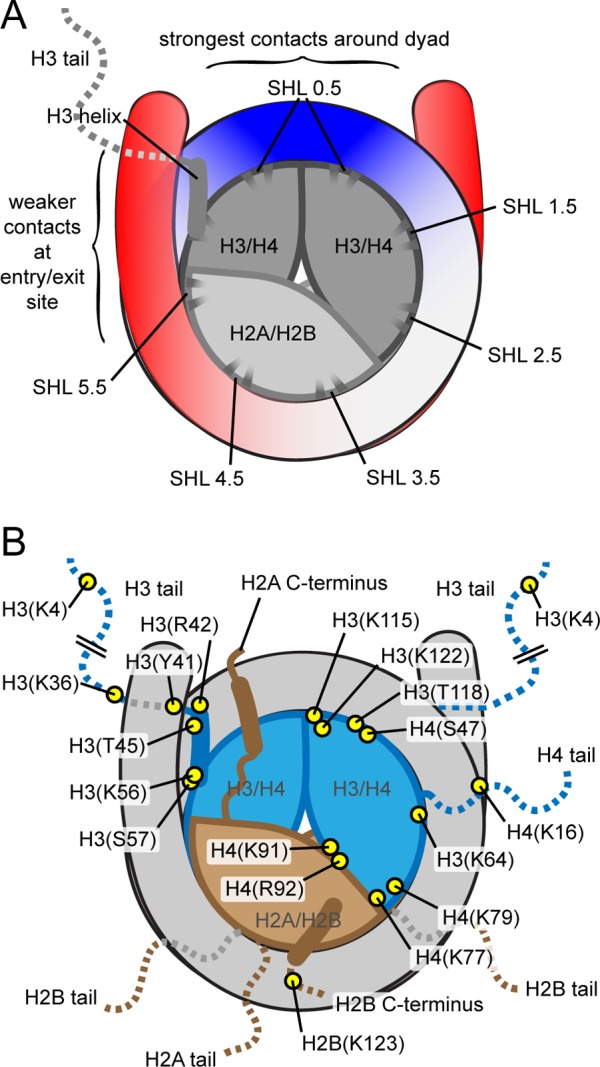
Schematic drawings of the nucleosome, highlighting
features that
contribute to nucleosome dynamics. (A) Illustration highlighting energetically
important contacts within the nucleosome. The DNA entry/exit region
(red) has weaker histone–DNA contacts, and the dyad region
(blue) has the most energetically important contacts. Superhelical locations (SHLs) indicate the histone surfaces
where contact is made with the DNA minor groove. The histone surface underneath the DNA at the dyad, where
the major groove faces the histone octamer, is considered SHL-0, and
increasing or decreasing values mark each SHL moving from the dyad
to the two entry/exit regions. (B) Map of histone residues where post-translational
modifications influence nucleosome dynamics.
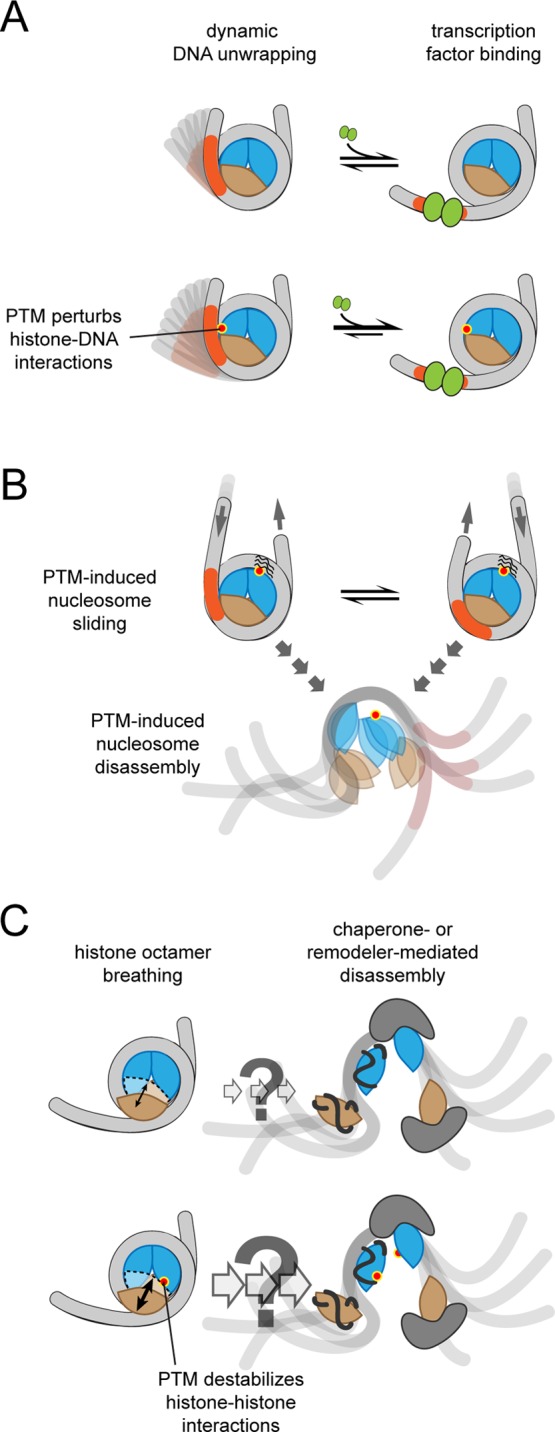
Types
of nucleosome dynamics that can be affected by PTMs. (A)
DNA unwrapping transiently exposes protein-binding sites that are
buried within the fully wrapped nucleosome. (B) The DNA can slide
relative to the histone octamer and nucleosomes can be disassembled
to expose DNA-binding sites. With unmodified histones, these structural
changes require histone chaperones and chromatin remodeling complexes.
Dyad modifications can enhance both sliding and disassembly. (C) Nucleosomes
can unwrap with the H2A/H2B heterodimer attached to DNA. This type
of structural dynamics could be an initial step for nucleosome disassembly
and H2A/H2B exchange and may be accelerated by PTMs at histone–histone
interfaces.
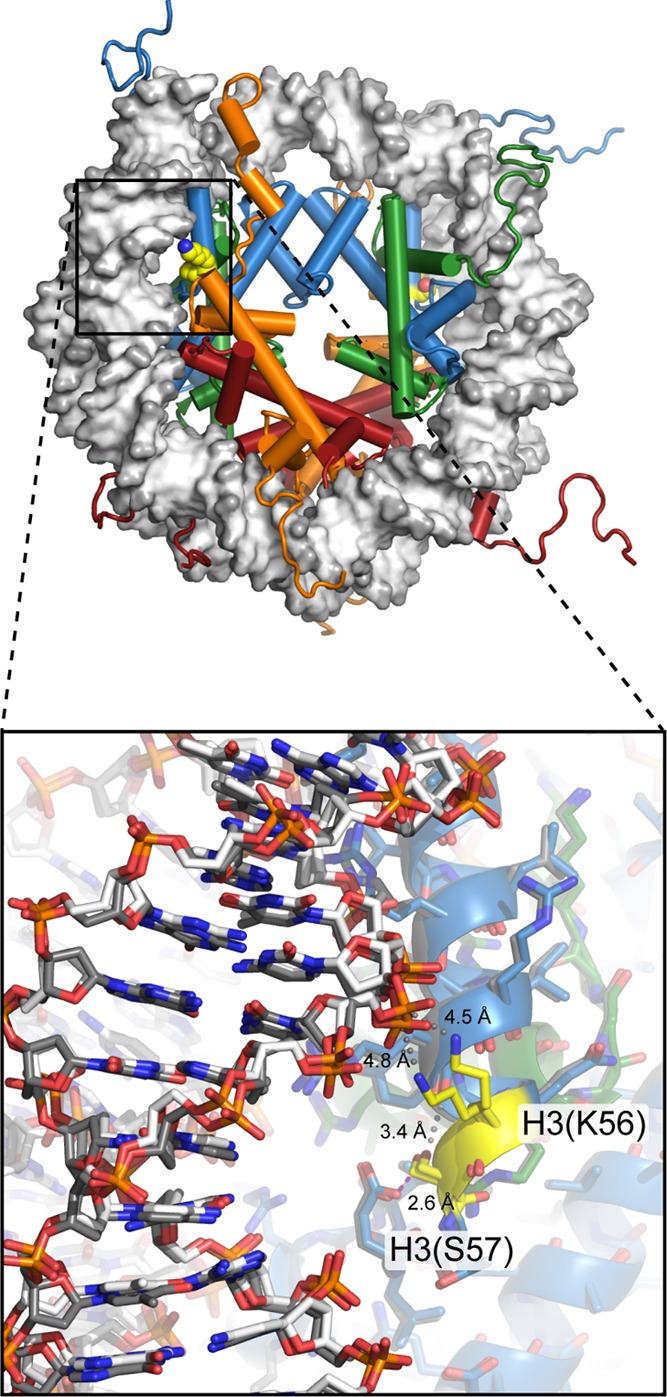
View of
the nucleosome (1KX5), with H3(K56) and H3(S57) highlighted in yellow.
Located under the DNA near the nucleosome entry/exit region, H3(K56ac)
increases site exposure by increasing the DNA unwrapping rate, and influences histone chaperone binding, while H3(S57A) substitution interferes with octamer formation and
increases H2A/H2B dimer exchange. Close-up
view (bottom) shows the two sides of the nucleosome superimposed,
with one copy of each histone in color and one copy in gray. In the
crystal structure, H3(S57) hydrogen bonds with neighboring H3(E59)
(magenta dotted line) and makes van der Waals contacts with carbonyl
oxygens of the αN-helix of histone H3 (gray spheres) but is
too far to make direct interactions with DNA. Although the neighboring
H3(K56) makes a closer approach to DNA, the lysine side chain is too
distant to directly hydrogen-bond to the phosphate backbone. The two
positions observed for the H3(K56) side chain suggest some mobility,
and the small gray spheres highlight the shortest path from the lysine
to the closest DNA phosphates.
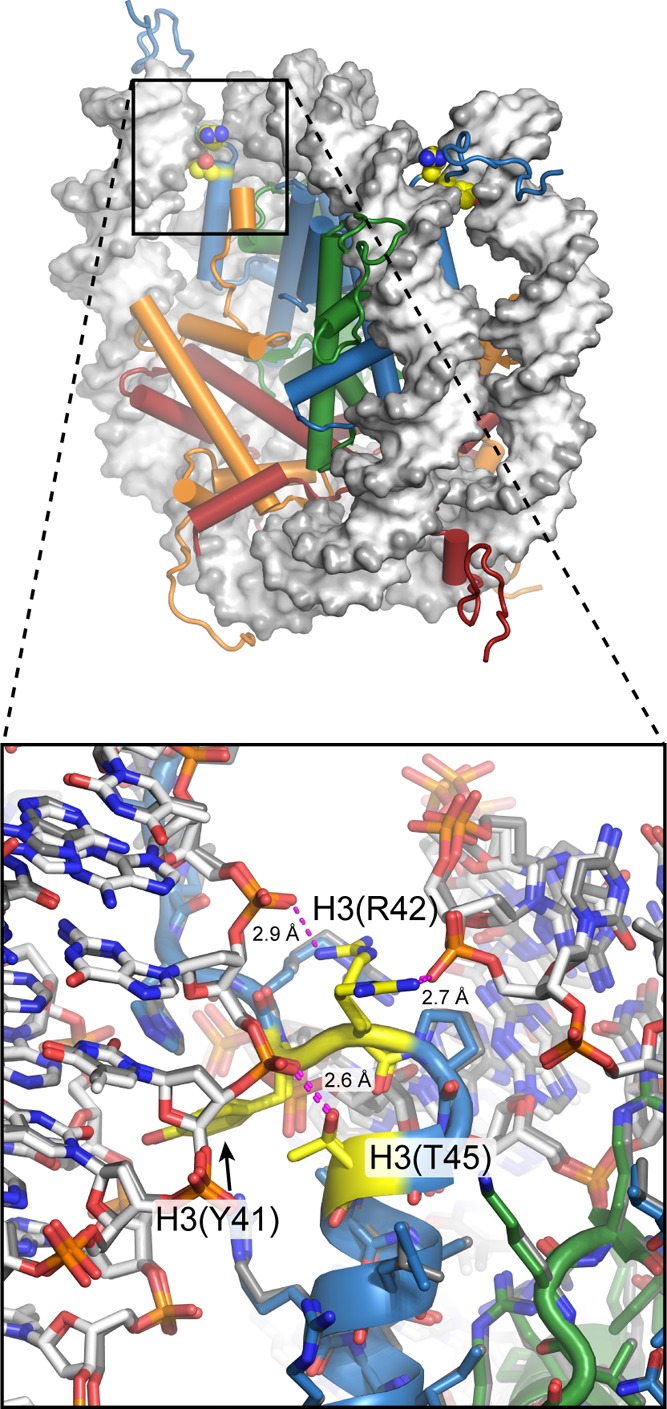
View of the nucleosome (1KX5), with H3(Y41), H3(R42), and H3(T45) highlighted in
yellow. These residues are located right where the DNA enters and
exits the nucleosome. Close-up view (bottom) with the two sides of
the nucleosome superimposed shows the same rotamers for H3(Y41) and
H3(T45) and two different conformations for H3(R42). In this structure,
both H3(R42) and H3(T45) make direct hydrogen bonds to the DNA phosphate
backbone (magenta dotted lines). Phosphorylation of Y41 or T45 would
be expected to cause steric clashes and electrostatic repulsion with
the DNA. The direct impact of PTMs at these positions on nucleosome
dynamics has yet to be reported, but they are predicted to increase
DNA unwrapping on the basis of studies of H3(K56ac).
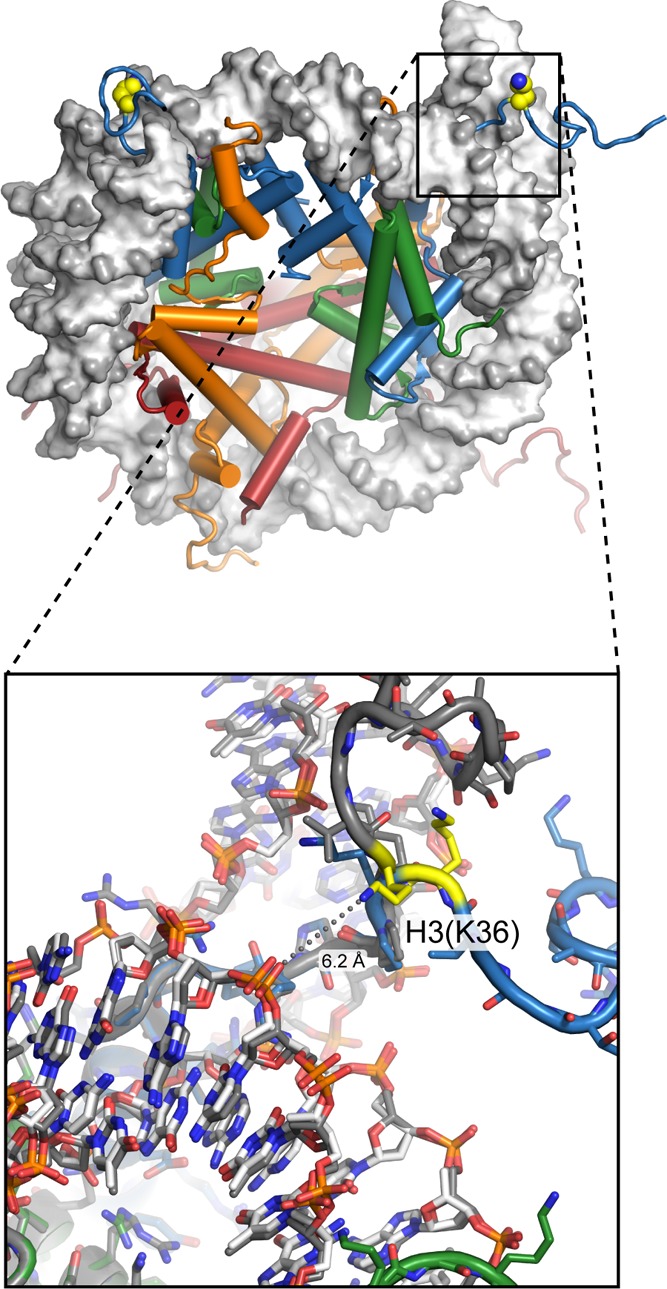
View of the nucleosome (1KX5), with H3(K36) highlighted
in yellow. This residue
is located on the H3 tail just outside where the tail enters between
the two gyres of DNA at the entry/exit region. Close-up view (bottom)
with the two sides of the nucleosome superimposed shows that even
the backbone position of H3(K36) differs between the two copies in
this structure. Neither copy of H3(K36) has the lysine side chain
within direct hydrogen-bonding distance of the DNA backbone. While
modification of this residue does not influence DNA unwrapping, the binding of the Phf1 Tudor domain enhances
DNA unwrapping and accessibility. This
suggests that other histone PTM readers that bind in the entry/exit
region may also alter nucleosome unwrapping/rewrapping dynamics.
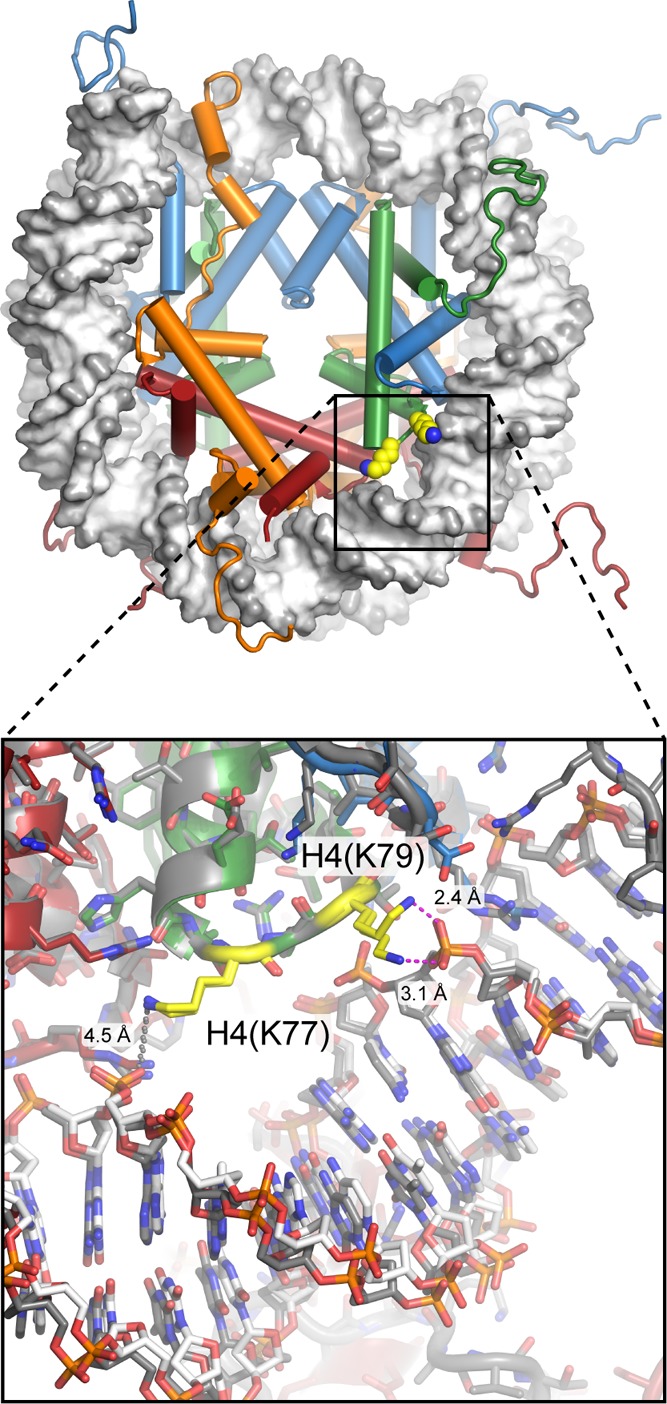
View of
the nucleosome (1KX5), with H4(K77) and H4(K79) highlighted in yellow.
These residues are located around SHL ± 3, where histone mutations
result in the loss of rDNA silencing (LRS). Close-up view (bottom) with the two sides of the nucleosome superimposed
shows that H4(K79) can occupy two different conformations yet still
directly hydrogen-bond to the DNA phosphate backbone and that H4(K77)
is too distant to directly hydrogen-bond in this structure. Despite
their location relatively far from the edge of the nucleosome, acetylation
of these two residues increases DNA unwrapping at the entry/exit site, which may underlie their connection to disrupting
transcriptional silencing.
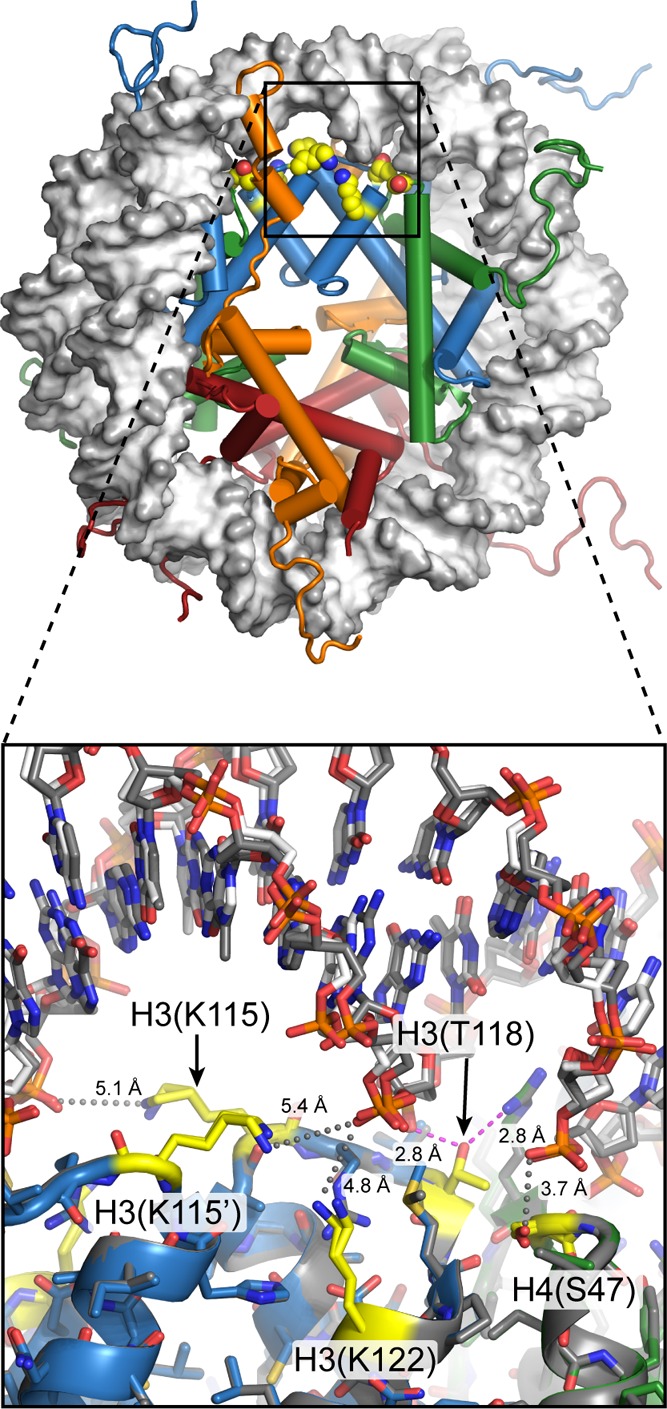
View of
the nucleosome (1KX5), with H3(K115), H3(T118), H3(K122), and H4(S47) highlighted
in yellow. Located around SHL ± 0.5, these residues are positioned
within the most energetically important histone–DNA contacts.
Close-up view (bottom) with the two sides of the nucleosome superimposed
shows very similar conformations for these residues. H3(T118) generates
a SIN phenotype when mutated and directly hydrogen-bonds to both the
DNA phosphate backbone and another SIN residue, H4(R45) (magenta dotted
lines). Phosphorylation of H3(T118), which would likely disrupt these
energetically important interactions, has been shown to destabilize
the nucleosome, similar to SIN mutations.,,
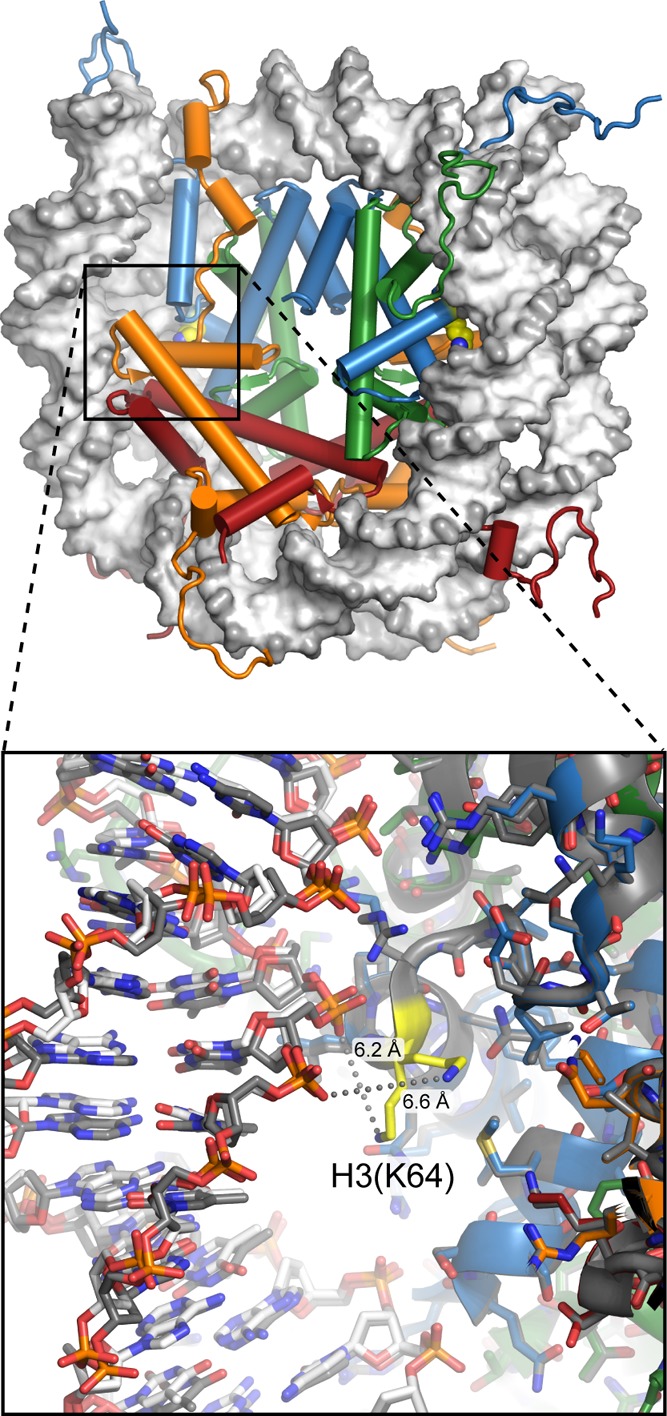
View of the nucleosome (1KX5), with H3(K64) shown
in yellow. Located at SHL ±
2, this position lies underneath the major groove of DNA and would
not be easily accessible to a histone-modifying enzyme in a fully
wrapped nucleosome. Close-up view (bottom) with the two sides of the
nucleosome superimposed shows very different positions of the lysine
side chain, neither within hydrogen-bonding distance of the DNA phosphate
backbone (gray spheres). From the crystal structure, the manner in
which modification of H3(K64) would directly impact nucleosome dynamics
is not clear.
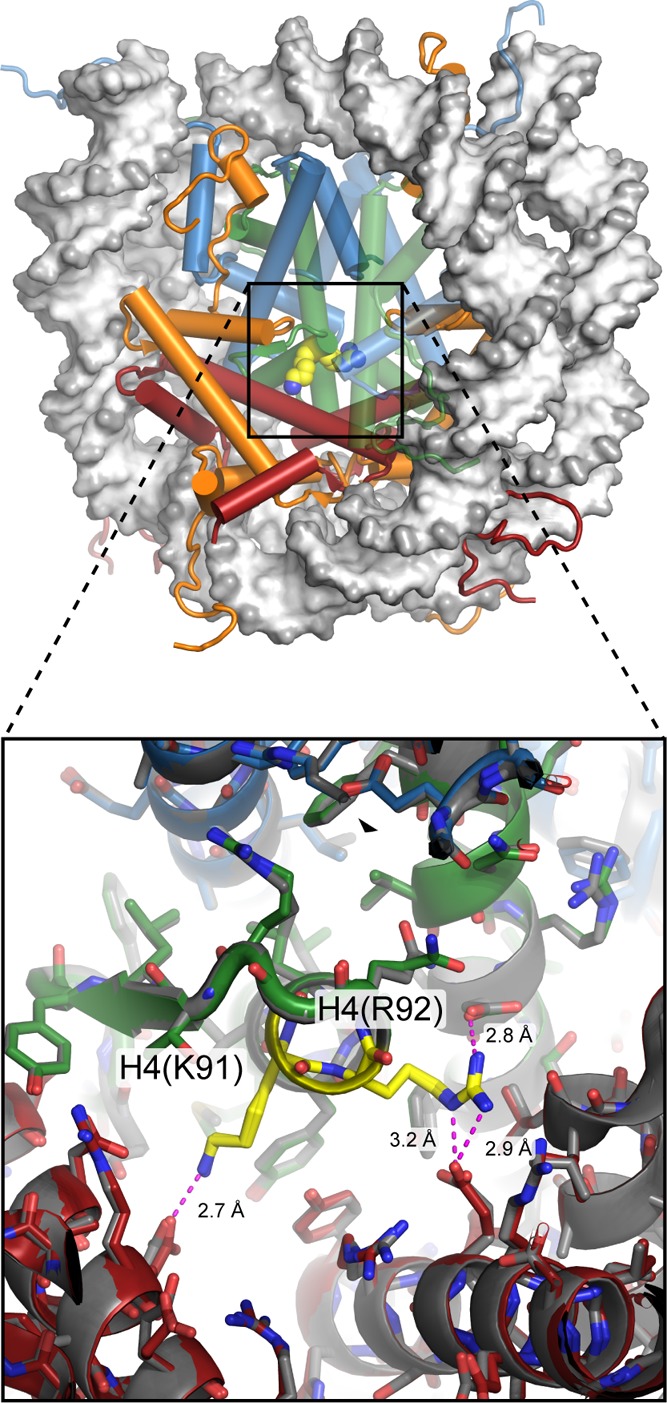
View of the nucleosome (1KX5), with H4(K91) and H4(R92) highlighted
in yellow.
These residues are located near the center of the histone octamer,
at the interface between H2A/H2B dimers and H3/H4 tetramer, and are
not readily accessible from the exterior of the nucleosome in the
crystal structure. In the far view (top), the backbone of H3/H4 on
the right side is semitransparent so that H4(R92) can be seen. Close-up
view (bottom) with the two sides of the nucleosome superimposed shows
that both copies are in very similar conformations, with each making
direct hydrogen bonds to residues on H2B (magenta dotted lines). Modification
of either H4(K91) or H4(R92) would be expected to interfere with these
hydrogen bonds. H4(K91ac) is involved in nucleosome assembly and may
increase fluctuations or dissociation of H2A/H2B on the nucleosome.
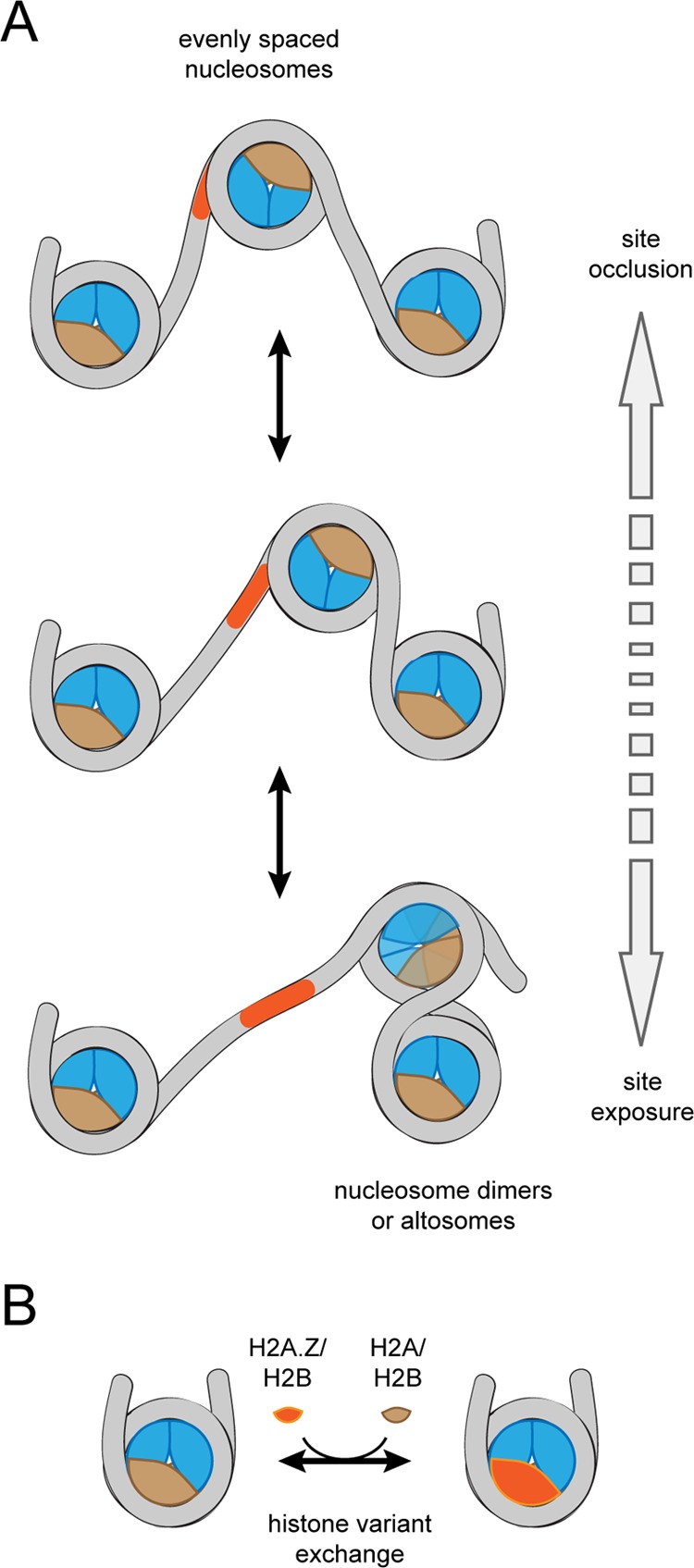
Changes in nucleosome organization carried out by chromatin remodelers.
(A) Most chromatin remodelers can reposition or “slide”
nucleosomes along DNA. Depending on the direction of sliding, this
repositioning can bury or expose DNA binding sites. Remodelers like
Chd1 and many ISWI-type remodelers are sensitive to DNA flanking the
nucleosome and generate evenly spaced nucleosome arrays (top). In
contrast, other remodelers such as SWI/SNF and RSC can shift nucleosomes
into their neighbors, generating dimeric or altosome structures, which
are believed to be intermediates for nucleosome disassembly (bottom).
The altosome organization depicted here was adapted from a model of
Ulyanova and Schnitzler. (B) Some remodelers
specialize in histone variant exchange. Exchange of canonical and
variant H2A/H2B dimers, highlighted here, is carried out by SWR1 and
INO80.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
