

Proton-mediated block of Ca2+ channels during multivesicular release regulates short-term plasticity at an auditory hair cell synapse
- PMID: 25429130
- PMCID: PMC4244462
- DOI: 10.1523/JNEUROSCI.2304-14.2014
Proton-mediated block of Ca2+ channels during multivesicular release regulates short-term plasticity at an auditory hair cell synapse
Abstract
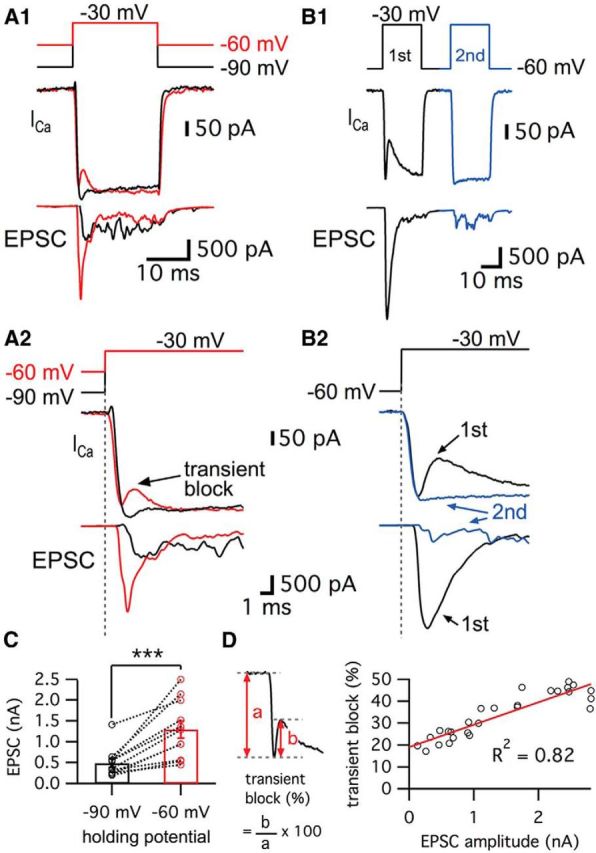
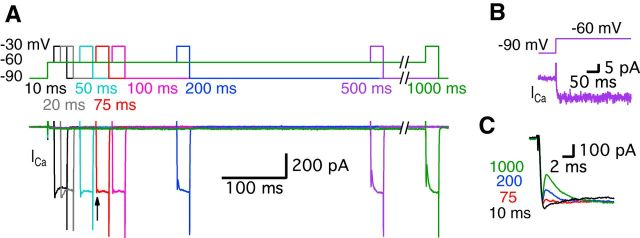
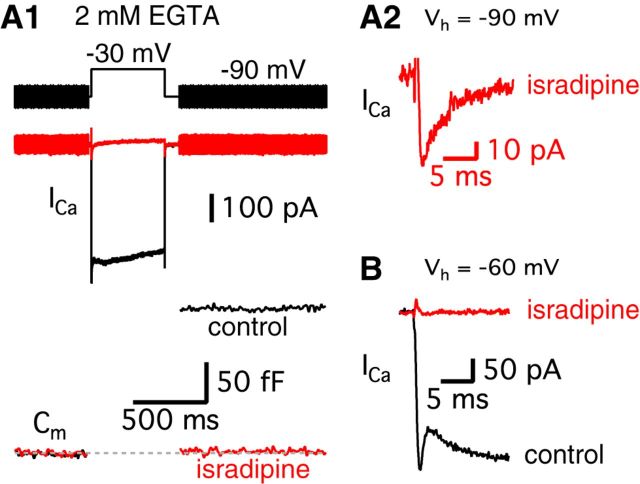
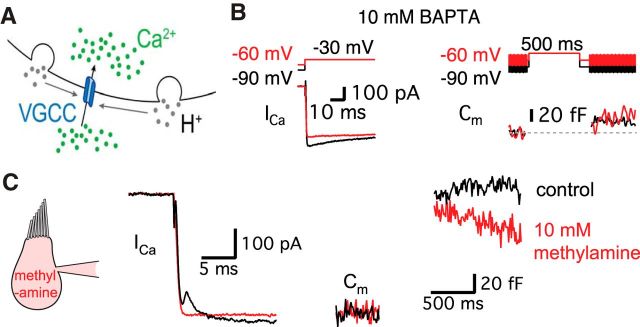
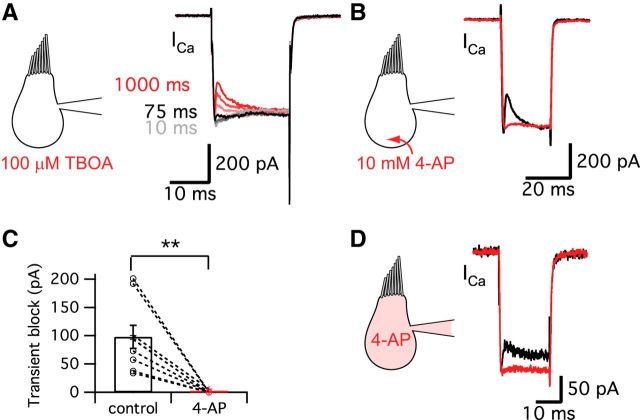
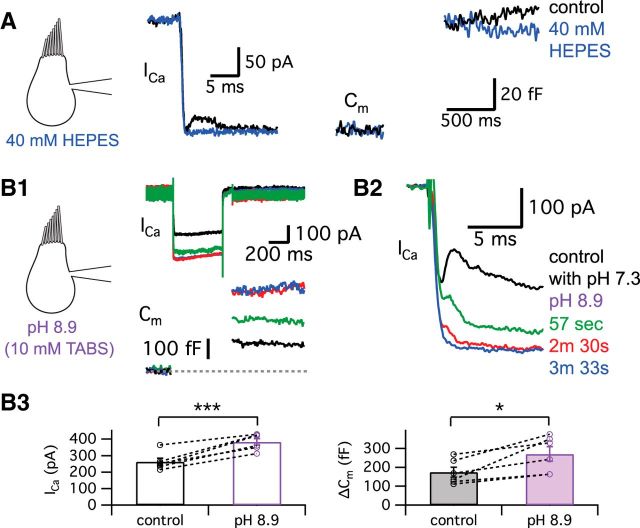
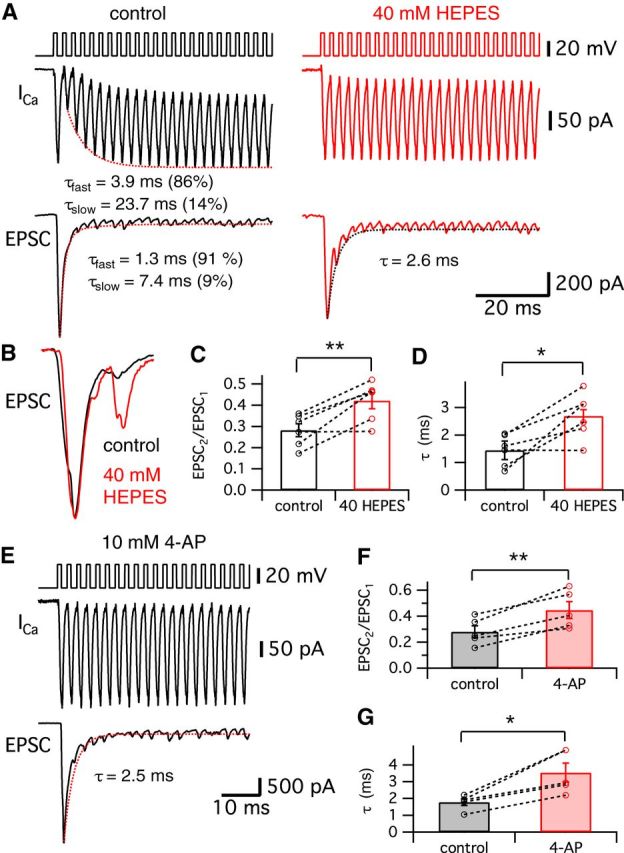
Synaptic vesicles release both neurotransmitter and protons during exocytosis, which may result in a transient acidification of the synaptic cleft that can block Ca(2+) channels located close to the sites of exocytosis. Evidence for this effect has been reported for retinal ribbon-type synapses, but not for hair cell ribbon synapses. Here, we report evidence for proton release from bullfrog auditory hair cells when they are held at more physiological, in vivo-like holding potentials (Vh = -60 mV) that facilitate multivesicular release. During paired recordings of hair cells and afferent fibers, L-type voltage-gated Ca(2+) currents showed a transient block, which was highly correlated with the EPSC amplitude (or the amount of glutamate release). This effect was masked at Vh = -90 mV due to the presence of a T-type Ca(2+) current and blocked by strong pH buffering with HEPES or TABS. Increasing vesicular pH with internal methylamine in hair cells also abolished the transient block. High concentrations of intracellular Ca(2+) buffer (10 mm BAPTA) greatly reduced exocytosis and abolished the transient block of the Ca(2+) current. We estimate that this transient block is due to the rapid multivesicular release of ∼600-1300 H(+) ions per synaptic ribbon. Finally, during a train of depolarizing pulses, paired pulse plasticity was significantly changed by using 40 mm HEPES in addition to bicarbonate buffer. We propose that this transient block of Ca(2+) current leads to more efficient exocytosis per Ca(2+) ion influx and it may contribute to spike adaptation at the auditory nerve.
Keywords: auditory; calcium current; electrophysiology; exocytosis; hair cells; protons.
Copyright © 2014 the authors 0270-6474/14/3415877-11$15.00/0.
Figures
References
-
- Ballanyi K, Kaila K. Activity-evoked changes in intracellular pH. In: Kaila K, Ransom BR, editors. pH and Brain Function. New York: Wiley; 1998. pp. 291–308.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous