

Abnormal emotional learning in a rat model of autism exposed to valproic acid in utero
- PMID: 25429264
- PMCID: PMC4228846
- DOI: 10.3389/fnbeh.2014.00387
Abnormal emotional learning in a rat model of autism exposed to valproic acid in utero
Abstract
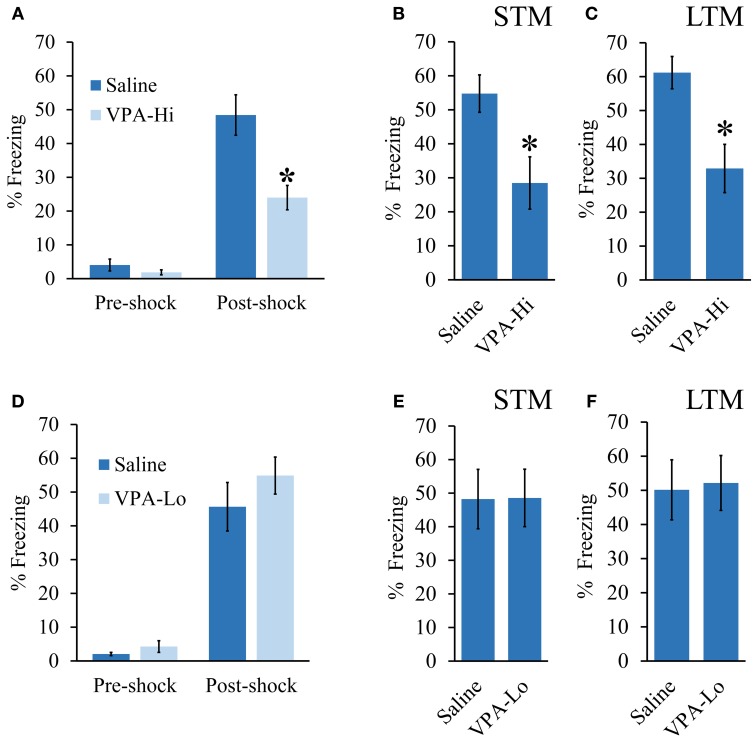
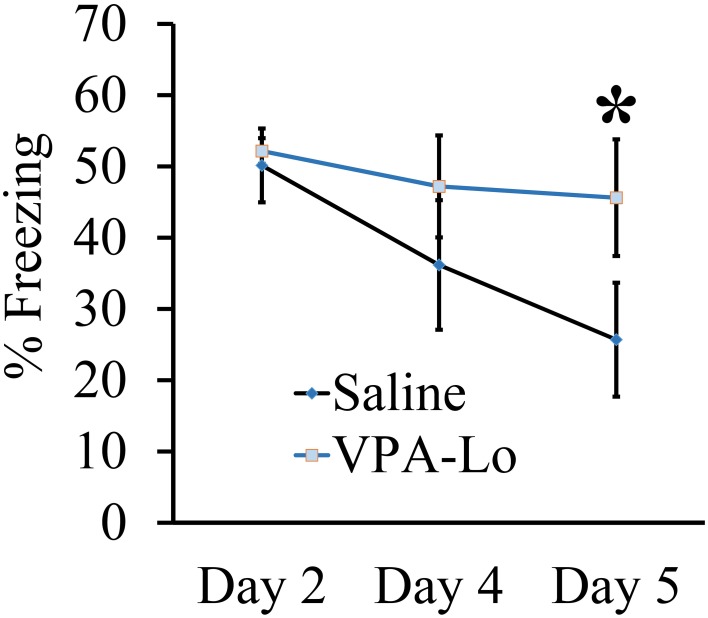
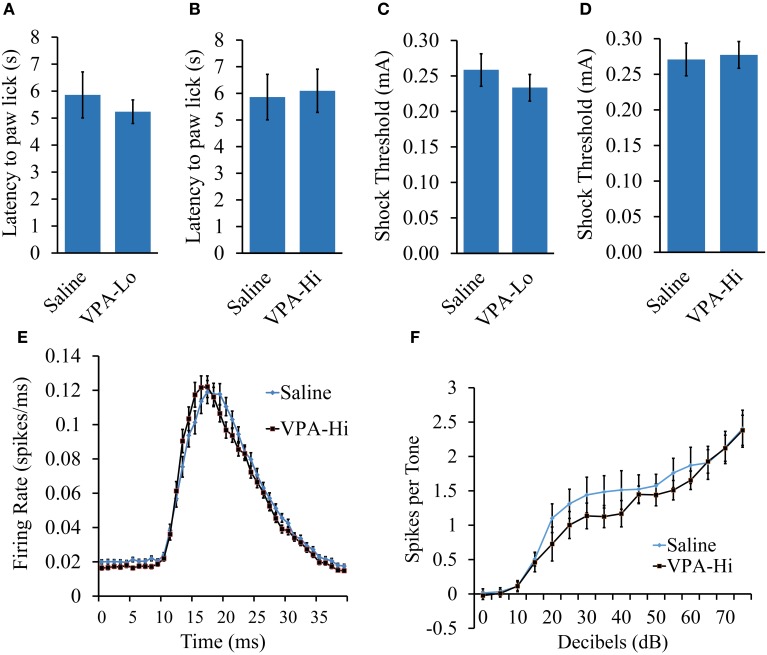
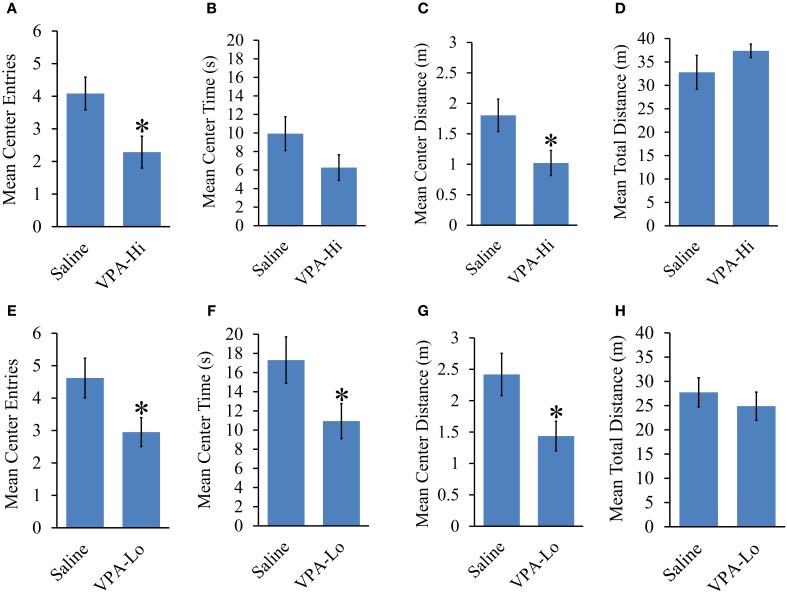
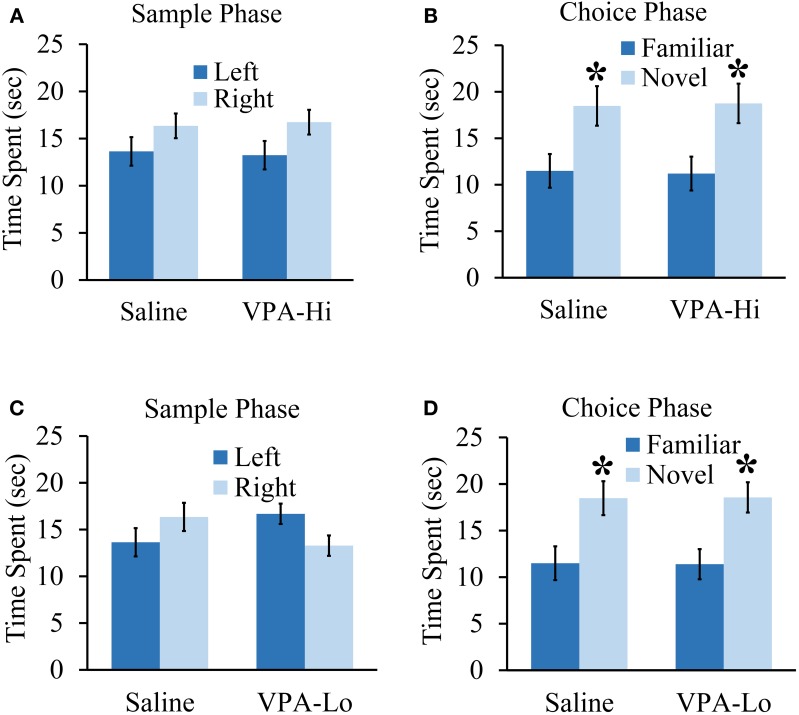
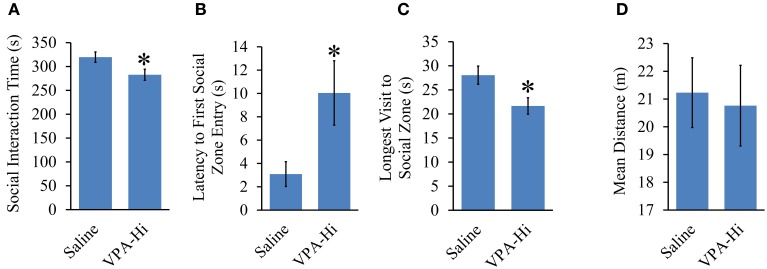
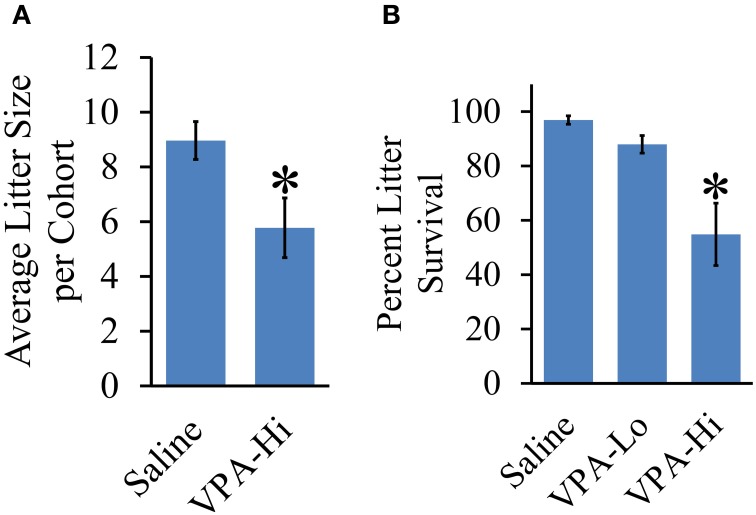
Autism Spectrum Disorders (ASD) are complex neurodevelopmental disorders characterized by repetitive behavior and impaired social communication and interactions. Apart from these core symptoms, a significant number of ASD individuals display higher levels of anxiety and some ASD individuals exhibit impaired emotional learning. We therefore sought to further examine anxiety and emotional learning in an environmentally induced animal model of ASD that utilizes the administration of the known teratogen, valproic acid (VPA) during gestation. Specifically we exposed dams to one of two different doses of VPA (500 and 600 mg/kg) or vehicle on day 12.5 of gestation and examined the resultant progeny. Our data indicate that animals exposed to VPA in utero exhibit enhanced anxiety in the open field test and normal object recognition memory compared to control animals. Animals exposed to 500 mg/kg of VPA displayed normal acquisition of auditory fear conditioning, and exhibited reduced extinction of fear memory and normal litter survival rates as compared to control animals. We observed that animals exposed to 600 mg/kg of VPA exhibited a significant reduction in the acquisition of fear conditioning, a significant reduction in social interaction and a significant reduction in litter survival rates as compared to control animals. VPA (600 mg/kg) exposed animals exhibited similar shock sensitivity and hearing as compared to control animals indicating the fear conditioning deficit observed in these animals was not likely due to sensory deficits, but rather due to deficits in learning or memory retrieval. In conclusion, considering that progeny from dams exposed to rather similar doses of VPA exhibit striking differences in emotional learning, the VPA model may serve as a useful tool to explore the molecular and cellular mechanisms that contribute to not only ASD, but also emotional learning.
Keywords: Pavlovian fear conditioning; amygdala; autism; emotion; learning; memory; valproic acid.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources