

Bioenergetic evolution in proteobacteria and mitochondria
- PMID: 25432941
- PMCID: PMC4986455
- DOI: 10.1093/gbe/evu257
Bioenergetic evolution in proteobacteria and mitochondria
Abstract
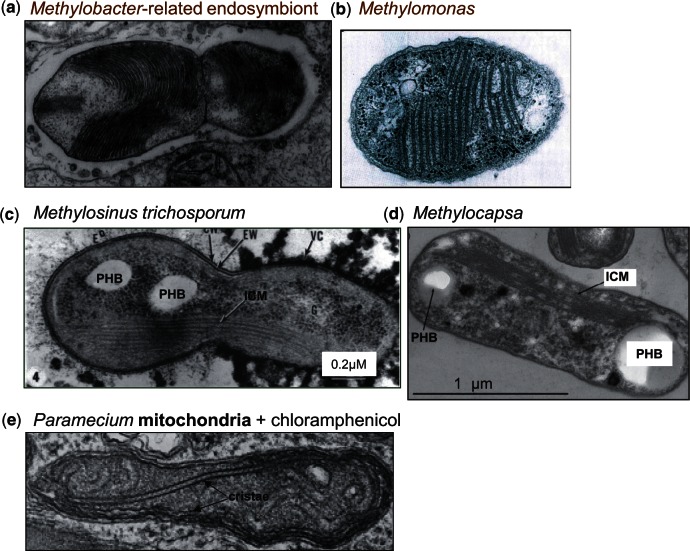
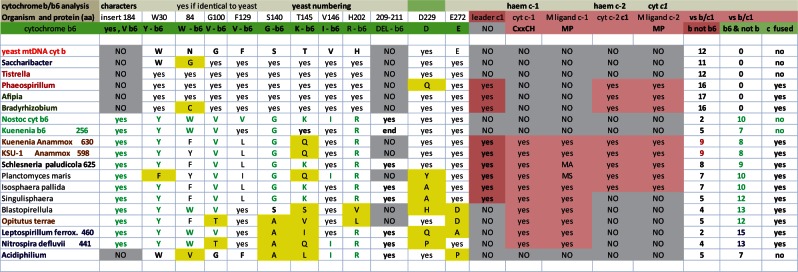
Mitochondria are the energy-producing organelles of our cells and derive from bacterial ancestors that became endosymbionts of microorganisms from a different lineage, together with which they formed eukaryotic cells. For a long time it has remained unclear from which bacteria mitochondria actually evolved, even if these organisms in all likelihood originated from the α lineage of proteobacteria. A recent article (Degli Esposti M, et al. 2014. Evolution of mitochondria reconstructed from the energy metabolism of living bacteria. PLoS One 9:e96566) has presented novel evidence indicating that methylotrophic bacteria could be among the closest living relatives of mitochondrial ancestors. Methylotrophs are ubiquitous bacteria that live on single carbon sources such as methanol and methane; in the latter case they are called methanotrophs. In this review, I examine their possible ancestry to mitochondria within a survey of the common features that can be found in the central and terminal bioenergetic systems of proteobacteria and mitochondria. I also discuss previously overlooked information on methanotrophic bacteria, in particular their intracytoplasmic membranes resembling mitochondrial cristae and their capacity of establishing endosymbiotic relationships with invertebrate animals and archaic plants. This information appears to sustain the new idea that mitochondrial ancestors could be related to extant methanotrophic proteobacteria, a possibility that the genomes of methanotrophic endosymbionts will hopefully clarify.
Keywords: bioenergetics; endosymbiosis; methanotrophs; methylotrophs; mitochondria.
© The Author(s) 2014. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
Similar articles
-
Evolution of mitochondria reconstructed from the energy metabolism of living bacteria.PLoS One. 2014 May 7;9(5):e96566. doi: 10.1371/journal.pone.0096566. eCollection 2014. PLoS One. 2014. PMID: 24804722 Free PMC article.
-
[Evolution of mitochondria].Tsitol Genet. 2002 Sep-Oct;36(5):49-57. Tsitol Genet. 2002. PMID: 12442548 Review. Russian.
-
The Origin of Mitochondrial Cristae from Alphaproteobacteria.Mol Biol Evol. 2017 Apr 1;34(4):943-956. doi: 10.1093/molbev/msw298. Mol Biol Evol. 2017. PMID: 28087774 Review.
-
[Suggested mitochondrial ancestry of non-mitochondrial ATP/ADP].Mol Biol (Mosk). 2007 Jan-Feb;41(1):59-70. Mol Biol (Mosk). 2007. PMID: 17380892 Russian.
-
Ancient homology of the mitochondrial contact site and cristae organizing system points to an endosymbiotic origin of mitochondrial cristae.Curr Biol. 2015 Jun 1;25(11):1489-95. doi: 10.1016/j.cub.2015.04.006. Epub 2015 May 21. Curr Biol. 2015. PMID: 26004762
Cited by
-
Division of labour in a matrix, rather than phagocytosis or endosymbiosis, as a route for the origin of eukaryotic cells.Biol Direct. 2020 Apr 28;15(1):8. doi: 10.1186/s13062-020-00260-9. Biol Direct. 2020. PMID: 32345370 Free PMC article.
-
Mitochondrial Dynamics during Development.Newborn (Clarksville). 2023 Jan-Mar;2(1):19-44. doi: 10.5005/jp-journals-11002-0053. Epub 2023 Apr 6. Newborn (Clarksville). 2023. PMID: 37206581 Free PMC article.
-
Oxygen Reductases in Alphaproteobacterial Genomes: Physiological Evolution From Low to High Oxygen Environments.Front Microbiol. 2019 Mar 18;10:499. doi: 10.3389/fmicb.2019.00499. eCollection 2019. Front Microbiol. 2019. PMID: 30936856 Free PMC article.
-
Edeine B1 produced by Brevibacillus brevis reduces the virulence of a plant pathogenic fungus by inhibiting mitochondrial respiration.mBio. 2024 Jul 17;15(7):e0135124. doi: 10.1128/mbio.01351-24. Epub 2024 Jun 11. mBio. 2024. PMID: 38860787 Free PMC article.
-
When Friendship Turns Sour: Effective Communication Between Mitochondria and Intracellular Organelles in Parkinson's Disease.Front Cell Dev Biol. 2020 Nov 30;8:607392. doi: 10.3389/fcell.2020.607392. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33330511 Free PMC article. Review.
References
-
- Abhishek A, Bavishi A, Bavishi A, Choudhary M. Bacterial genome chimaerism and the origin of mitochondria. Can J Microbiol. 2011;57:49–61. - PubMed
-
- Andersson SG, Zomorodipour A, et al. The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature. 1998;396:133–140. - PubMed
-
- Atteia A, et al. A proteomic survey of Chlamydomonas reinhardtii mitochondria sheds new light on the metabolic plasticity of the organelle and on the nature of the alpha-proteobacterial mitochondrial ancestor. Mol Biol Evol. 2009;26:1533–1548. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
