

Short-chain fatty acid sensing in rat duodenum
- PMID: 25433076
- PMCID: PMC4324707
- DOI: 10.1113/jphysiol.2014.280792
Short-chain fatty acid sensing in rat duodenum
Abstract
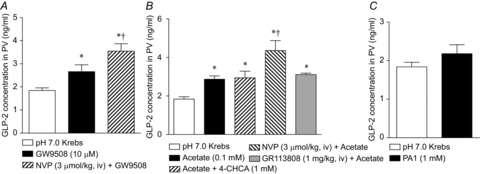
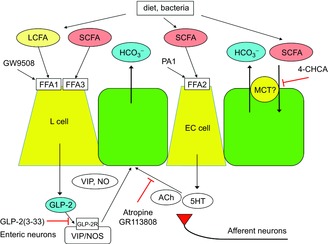
Key points: Luminal lipid in the duodenum modulates gastroduodenal functions via the release of gut hormones and mediators such as cholecystokinin and 5-HT. The effects of luminal short-chain fatty acids (SCFAs) in the foregut are unknown. Free fatty acid receptors (FFARs) for long-chain fatty acids (LCFAs) and SCFAs are expressed in enteroendocrine cells. SCFA receptors, termed FFA2 and FFA3, are expressed in duodenal enterochromaffin cells and L cells, respectively. Activation of LCFA receptor (FFA1) and presumed FFA3 stimulates duodenal HCO3(-) secretion via a glucagon-like peptide (GLP)-2 pathway, whereas FFA2 activation induces HCO3(-) secretion via muscarinic and 5-HT4 receptor activation. The presence of SCFA sensing in the duodenum with GLP-2 and 5-HT signals further supports the hypothesis that luminal SCFA in the foregut may contribute towards the generation of functional symptoms.
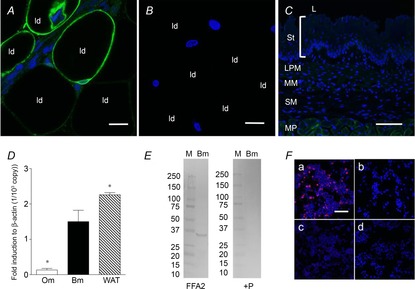
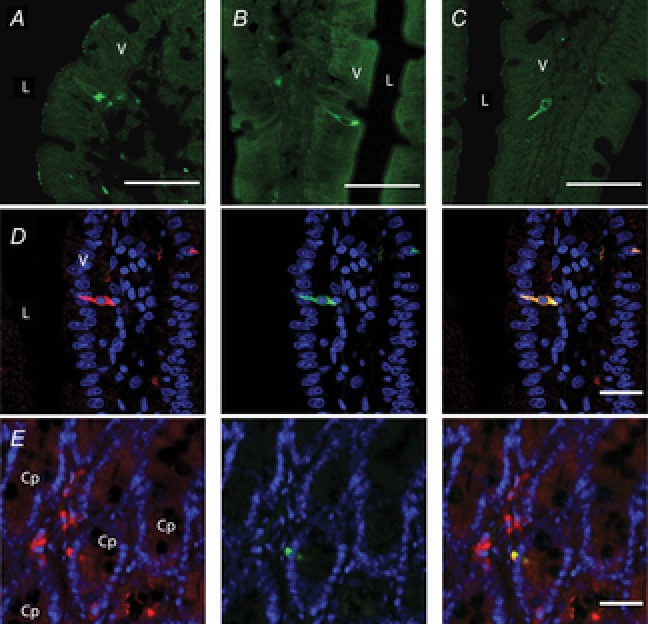
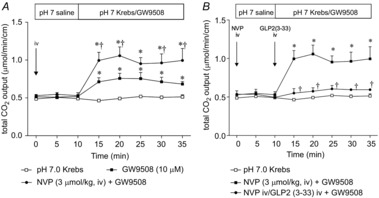
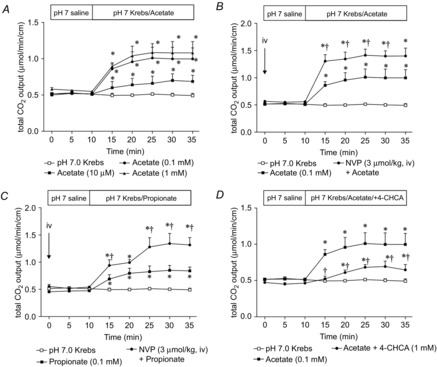
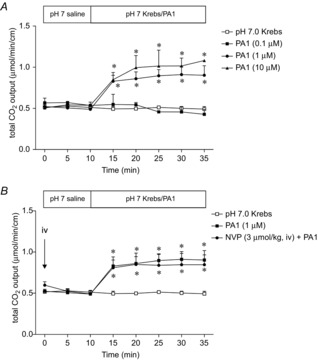
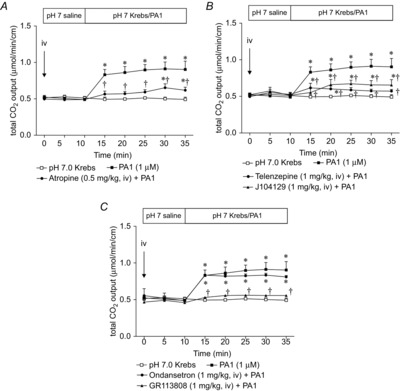
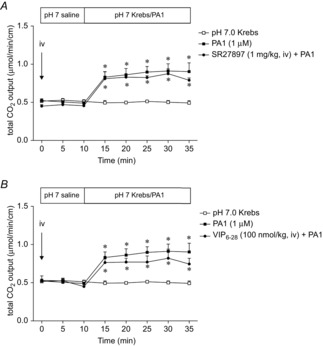
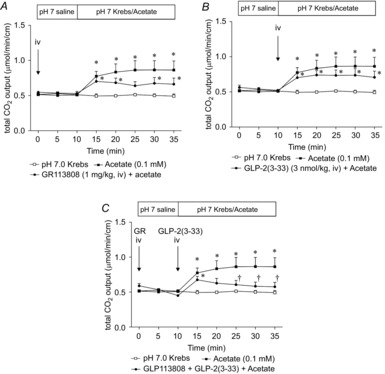
Abstract: Intraduodenal fatty acids (FA) and bacterial overgrowth, which generate short-chain FAs (SCFAs), have been implicated in the generation of functional dyspepsia symptoms. We studied the mechanisms by which luminal SCFA perfusion affects duodenal HCO3(-) secretion (DBS), a measure of mucosal neurohumoral activation. Free fatty acid receptor (FFAR) 1 (FFA1), which binds long-chain FA (LCFA), and SCFA receptors FFA2 and FFA3 were immunolocalised to duodenal enteroendocrine cells. FFA3 colocalised with glucagon-like peptide (GLP)-1, whereas FFA2 colocalised with 5-HT. Luminal perfusion of the SCFA acetate or propionate increased DBS, enhanced by dipeptidyl peptidase-IV (DPPIV) inhibition, at the same time as increasing GLP-2 portal blood concentrations. Acetate-induced DBS was partially inhibited by monocarboxylate/HCO3(-) exchanger inhibition without affecting GLP-2 release, implicating acetate absorption in the partial mediation of DBS. A selective FFA2 agonist dose-dependently increased DBS, unaffected by DPPIV inhibition or by cholecystokinin or 5-HT3 receptor antagonists, but was inhibited by atropine and a 5-HT4 antagonist. By contrast, a selective FFA1 agonist increased DBS accompanied by GLP-2 release, enhanced by DPPIV inhibition and inhibited by a GLP-2 receptor antagonist. Activation of FFA1 by LCFA and presumably FFA3 by SCFA increased DBS via GLP-2 release, whereas FFA2 activation stimulated DBS via muscarinic and 5-HT4 receptor activation. SCFA/HCO3(-) exchange also appears to be present in the duodenum. The presence of duodenal fatty acid sensing receptors that signal hormone release and possibly signal neural activation may be implicated in the pathogenesis of functional dyspepsia.
© 2014 The Authors. The Journal of Physiology © 2014 The Physiological Society.
Figures
References
-
- Akiba Y. Kaunitz JD. Duodenal chemosensing: Master control for epigastric sensation. J Gastroenterol Hepatol. 2011;26:6–7. - PubMed
-
- Akiba Y, Mizumori M, Guth PH, Engel E. Kaunitz JD. Duodenal brush border intestinal alkaline phosphatase activity affects bicarbonate secretion in rats. Am J Physiol Gastrointest Liver Physiol. 2007;293:G1223–G1233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous