

From in vitro to in vivo Models of Bacterial Biofilm-Related Infections
- PMID: 25437038
- PMCID: PMC4235718
- DOI: 10.3390/pathogens2020288
From in vitro to in vivo Models of Bacterial Biofilm-Related Infections
Abstract
The influence of microorganisms growing as sessile communities in a large number of human infections has been extensively studied and recognized for 30-40 years, therefore warranting intense scientific and medical research. Nonetheless, mimicking the biofilm-life style of bacteria and biofilm-related infections has been an arduous task. Models used to study biofilms range from simple in vitro to complex in vivo models of tissues or device-related infections. These different models have progressively contributed to the current knowledge of biofilm physiology within the host context. While far from a complete understanding of the multiple elements controlling the dynamic interactions between the host and biofilms, we are nowadays witnessing the emergence of promising preventive or curative strategies to fight biofilm-related infections. This review undertakes a comprehensive analysis of the literature from a historic perspective commenting on the contribution of the different models and discussing future venues and new approaches that can be merged with more traditional techniques in order to model biofilm-infections and efficiently fight them.
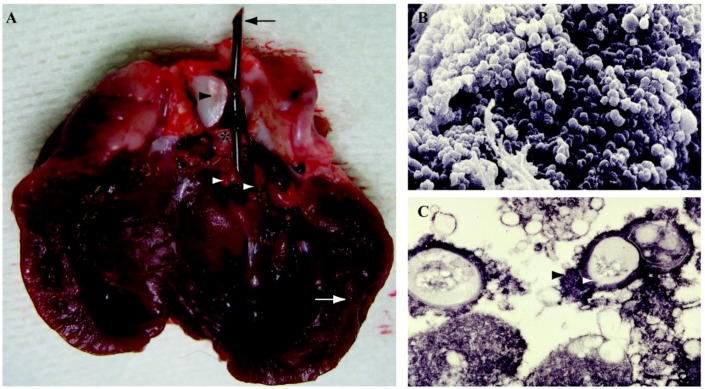
Figures





References
-
- Costerton J.W., Geesey G.G., Cheng K.J. How bacteria stick. Sci. Am. 1978;238:86–95. - PubMed
-
- Geesey G.G., Richardson W.T., Yeomans H.G., Irvin R.T., Costerton J.W. Microscopic examination of natural sessile bacterial populations from an alpine stream. Can. J. Microbiol. 1977;23:1733–1736. - PubMed
-
- Al-Azemi A., Fielder M.D., Abuknesha R.A., Price R.G. Effects of chelating agent and environmental stresses on microbial biofilms: relevance to clinical microbiology. J. Appl. Microbiol. 2011;110:1307–1313. - PubMed
-
- Hoiby N., Bjarnsholt T., Givskov M., Molin S., Ciofu O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents. 2010;35:322–332. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
