

Sphingolipid metabolism regulates development and lifespan in Caenorhabditis elegans
- PMID: 25437839
- PMCID: PMC4292899
- DOI: 10.1016/j.mad.2014.11.002
Sphingolipid metabolism regulates development and lifespan in Caenorhabditis elegans
Abstract
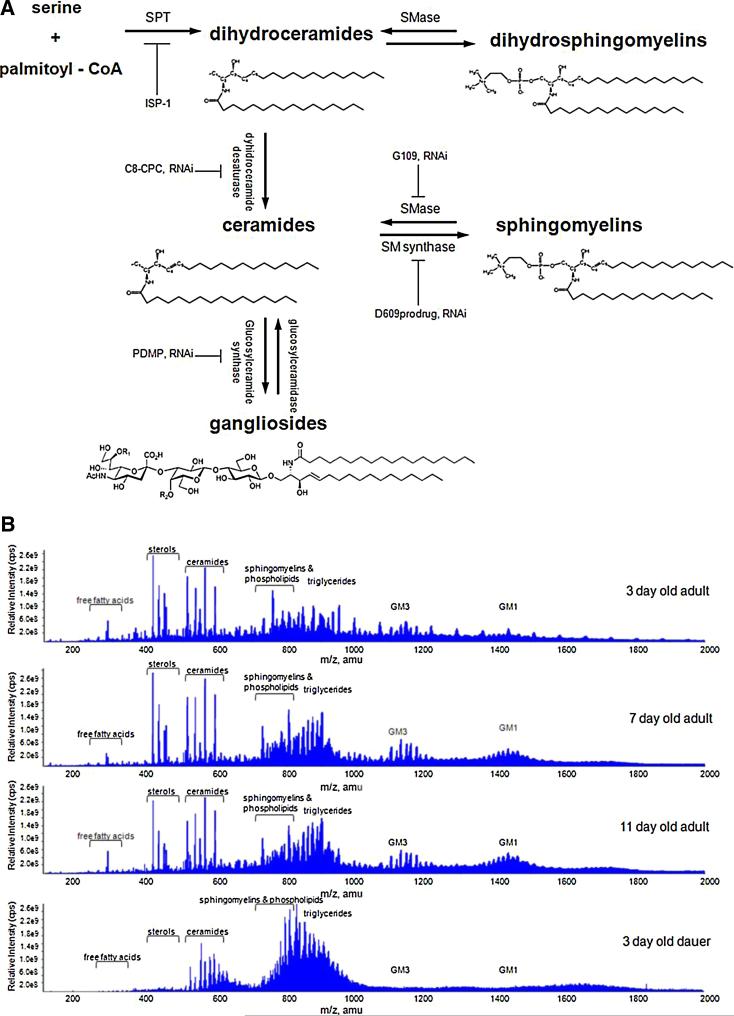
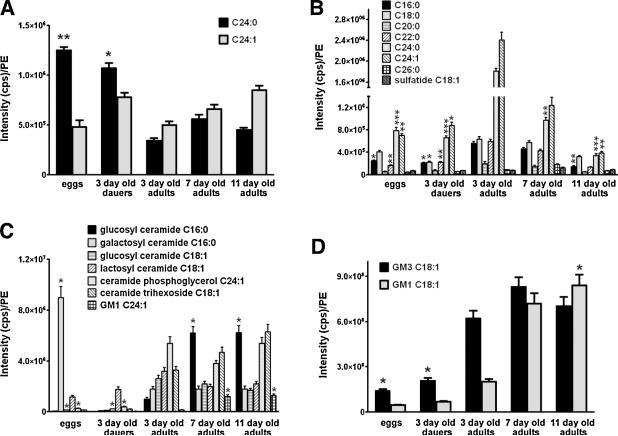
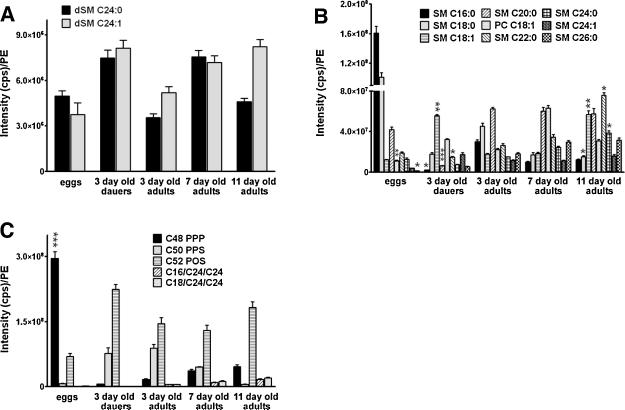
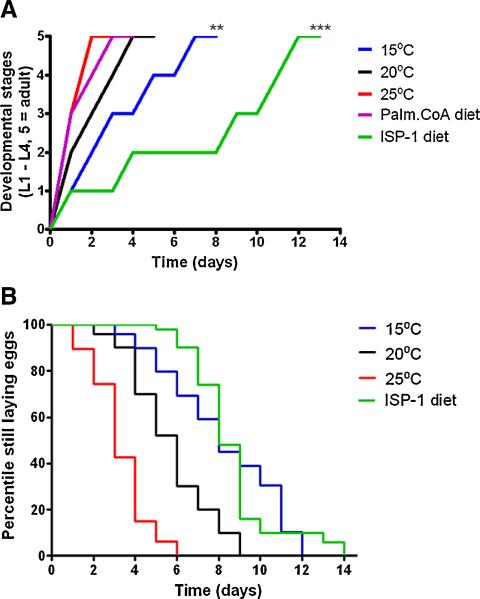
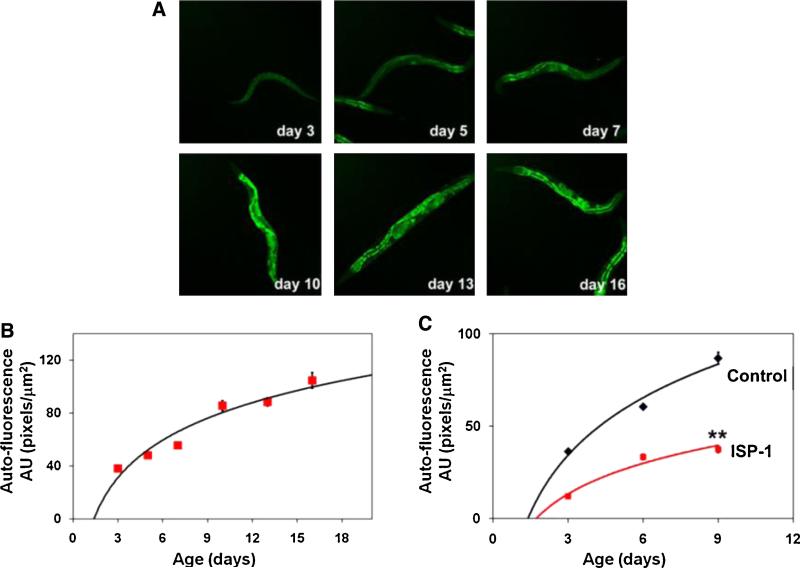
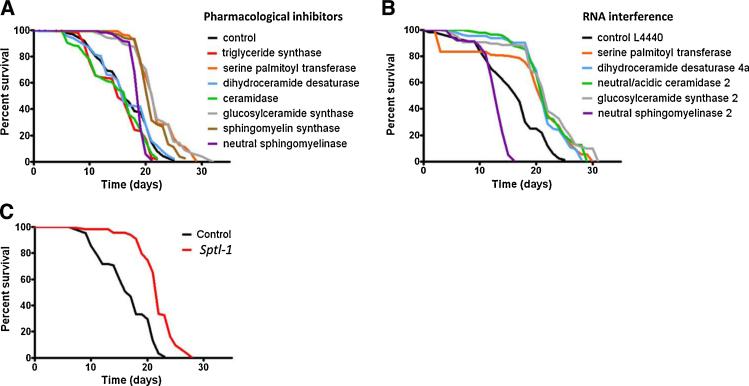
Sphingolipids are a highly conserved lipid component of cell membranes involved in the formation of lipid raft domains that house many of the receptors and cell-to-cell signaling factors involved in regulating cell division, maturation, and terminal differentiation. By measuring and manipulating sphingolipid metabolism using pharmacological and genetic tools in Caenorhabditis elegans, we provide evidence that the synthesis and remodeling of specific ceramides (e.g., dC18:1-C24:1), gangliosides (e.g., GM1-C24:1), and sphingomyelins (e.g., dC18:1-C18:1) influence development rate and lifespan. We found that the levels of fatty acid chain desaturation and elongation in many sphingolipid species increased during development and aging, with no such changes in developmentally-arrested dauer larvae or normal adults after food withdrawal (an anti-aging intervention). Pharmacological inhibitors and small interfering RNAs directed against serine palmitoyl transferase and glucosylceramide synthase acted to slow development rate, extend the reproductive period, and increase lifespan. In contrast, worms fed an egg yolk diet rich in sphingolipids exhibited accelerated development and reduced lifespan. Our findings demonstrate that sphingolipid accumulation and remodeling are critical events that determine development rate and lifespan in the nematode model, with both development rate and aging being accelerated by the synthesis of sphingomyelin, and its metabolism to ceramides and gangliosides.
Keywords: Ceramide; Gangliosides; Longevity; Oxidative stress; Sphingomyelin.
Published by Elsevier Ireland Ltd.
Figures
References
-
- Bielawska A, Greenberg MS, Perry D, Jayadev S, Shayman JA, McKay C, Hannun YA. (1S, 2R)-D-erythro-2-(N-myristoylamino)-1-phenyl-1-propanol as an inhibitor of ceramidase. J. Biol. Chem. 1996;271:12646–12654. - PubMed
-
- Bobryshev YV, Golovanova NK, Tra D, Samovilova NN, Gracheva EV, Efremov EE, Sobolev AY, Yurchenko YV, Lor RS, Cao W, Lu J, Saito M, Prokazova NV. Expression of GM3 synthase in human atherosclerotic lesions. Atherosclerosis. 2006;184:63–71. - PubMed
-
- Cutler RG, Pedersen WA, Camandola S, Rothstein JD, Mattson MP. Evidence that accumulation of ceramides and cholesterol esters mediates oxidative stress-induced death of motor neurons in amyotrophic lateral sclerosis. Ann. Neurol. 2002;52:448–457. - PubMed
-
- Cutler RG, Haughey NJ, Tammara A, McArthur JC, Nath A, Reid R, Vargas DL, Pardo CA, Mattson MP. Dysregulation of sphingolipid and sterol metabolism by ApoE4 in HIV dementia. Neurology. 2004;63:626–630. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
